I am organizing the colloquium for the Stanford Anthropology department this winter. I believe it may be the first time that a faculty member for the Ecology and Environment group has organized the colloquium since the Blessed Event that merged departments back in 2008 (though I'm not certain of that). There have been a few scheduling glitches, as it seems winter quarter 2015 has the highest density of talks I've yet encountered in 11 years at Stanford, but we're off to a great start. Our first speaker came all the way from the UK to speak to us about social dilemmas and cooperation. Shakti Lamba is an ESRC Research Fellow and Lecturer in Human Behavioural Ecology in the Centre for Ecology and Conservation at the University of Exeter.
Shakti talked about her very exciting work on behavioral norms. She uses a variety of methods, including ethnography, experimental games, and advanced statistical techniques to understand the nature of variation in cooperative norms within and between populations (see, e.g., papers here or here for examples of her work). I generally have mixed feelings about experimental games, but I think there is a small cadre of anthropologists, including Shakti and Drew Gerkey, among others, who use them as a tool for eliciting much richer behavioral and social observations than do most field researchers (whether or not they use experimental games!). I was impressed by the sophistication of her approach, her keen experimental design, and the excellent population thinking that it entails. However, I was most impressed with her coolness and eloquence under some pretty heated questioning from a number of senior faculty members who simply misunderstand evolutionary process. Looking forward to seeing more of her work, especially forthcoming longitudinal research with Alex Alvergne, in the future!
I just read an excellent piece at Slate.com this morning by Benjamin Hale. He notes that the scariest, most insidious thing about Ebola Virus Disease is that the disease capitalizes on intimate contact for transmission. While diseases such as influenza or cholera are transmitted by casual contact, frequently to strangers, via aerosolized droplets (influenza) or fecally contaminated water (cholera). Caretakers, and especially women, are hit hard by EVD. Hale writes,
...the mechanism Ebola exploits is far more insidious. This virus preys on care and love, piggybacking on the deepest, most distinctively human virtues. Affected parties are almost all medical professionals and family members, snared by Ebola while in the business of caring for their fellow humans. More strikingly, 75 percent of Ebola victims are women, people who do much of the care work throughout Africa and the rest of the world. In short, Ebola parasitizes our humanity.
True, and tragic, enough. But this article falls prey to one of my biggest frustrations with the reporting of science, one that I have written about recently in the context of the current EVD epidemic ravaging West Africa.
In the list Hale presents of the major concerns about EVD, he notes: "The threat of mutation," citing concern that Ebola virus might become airborne in a news report in Nature and the New York Times article that got me so worked up 10 days ago. Earlier this week, there was yet another longish piece in Nature/Scientific American that mentions "mutation" seven times but never once mentions selection. Or in another Nature piece, UCSF infectious disease physician Charles Chiu is quoted: "The longer we allow the outbreak to continue, the greater the opportunity the virus has to mutate, and it’s possible that it will mutate into a form that would be an even greater threat than it is right now.” True, mutations accumulate over time. Not true, mutation alone will make Ebola virus a greater threat than it is now. That would require selection.
While the idea of airborne transmission of Ebola virus is terrifying, the development of the ability to be transmitted via droplet or aerosol would be an adaptation on the part of the virus. Adaptations arise from the action of selection on the phenotypic variation. Phenotypes with higher fitness come to dominate the population of entities of which they are a part. In the case of a virus such as Ebola virus, this means that the virus must make sufficient copies of itself to ensure transmission to new susceptible hosts before killing the current host or being cleared by the host's immune system. While efficient transmission of EVD by aerosol or droplet would be horrible, equally horrible would be an adaptation that allowed it to transmit more efficiently from a dead host. It's not entirely clear how long Ebola virus can persist in its infectious state in the environment. In a study designed to maximize its persistence (indoors, in the dark, under laboratory conditions), Sagripanti and colleagues found that Ebola virus can persist for six days. Under field conditions, it's probably much shorter, but CDC suggests that 24 hours in a reasonably conservative estimate.
The lack of a strong relationship between host survival and pathogen transmission is why cholera can be so devastatingly pathogenic. The cholera patient can produce 10-20 liters of diarrhea (known as "rice water stools") per day. These stools contain billions of Vibrio cholerae bacteria, which enter the water supply and can infect other people at a distance well after the original host has died. The breaking of the trade-off between host mortality and the transmissibility of the pathogen means that the natural break on virulence is removed and the case fatality ratio can exceed 50%. That's high, kind of like the current round of EVD. Imagine if the trade-off between mortality and transmission in EVD were completely broken...
Changes in pathogen life histories like increased (or decreased) virulence or mode of transmission arise because of selection, not mutation, and this selection results from interactions with an environment that we are actively shaping. Sure, mutation matters because it provides raw material upon which selection can act, but the fact remains that we are talking primarily about selection here. Is this pervasive misunderstanding of the mechanisms of life the result of the war of misinformation being waged on science education in the US? I can't help but think it must at least be a contributor, but if it's true, it's pretty depressing because this misunderstanding is finding its way to some of the world's top news and opinion outlets.
This has to be a quick one, but I wanted to go on the record is noting my frustration at the current concern that Ebola might "mutate" into something far worse, like a pathogen that is efficiently transmitted by aerosol. For example, Michael Osterholm wrote in the New York Times yesterday, "The second possibility is one that virologists are loath to discuss openly but are definitely considering in private: that an Ebola virus could mutate to become transmissible through the air." I heard Morning Edition host David Greene ask WHO Director Margaret Chan last week, "Is this virus mutating in a way that could be very dangerous, that could make it spread faster?"
I agree, Ebola Virus becoming more easily transmitted by casual contact would be a 'nightmare scenario.' However, what we need to worry about is not mutation per se, but selection! Yes, the virus is mutating. It's a thing that viruses do. Ebola Virus is a Filovirus. It is composed of a single strand of negative-sense RNA. Like other viruses, and particularly RNA viruses, it is prone to high mutation rates. This is exacerbated by the fact that RNA polymerases lack the ability to correct mistakes. So mutations happen fast and they don't get cleaned up. Viruses also have very short generation times and can produce prodigious copies of themselves. This means that there is lots of raw material on which selection can act, because variation is the foundation of selection. Add to that heritability, which pretty much goes without saying since we are talking about the raw material of genetic information here, and differential transmission success and voilà, selection!
And virulence certainly responds to selection. There is a large literature on experimental evolution of virulence. See for example the many citations at the linked to Ebert's (1998) review in Sciencehere. There are lots of different specific factors that can favor the evolution of greater or lesser virulence and this is where theoretical biology can come in and make some sense of things. Steve Frank wrote a terrific review paper in 1996, available on his website, that describes many different models for the evolution of virulence. Two interesting regularities in the the evolution of virulence may be relevant to the current outbreak of EVD in West Africa. The first comes from a model developed by van Baalen & Sabelis (1995). Noting that there is an inherent trade-off between transmissibility of a pathogen and the extent of disease-induced mortality that it causes (a virus that makes more copies of itself is more likely to be transmitted but more viral copies means the host is sicker and might die), they demonstrate that when the relative transmissibility of a pathogen declines, its virulence will increase. They present a marginal value theorem solution for optimal virulence, which we can represent graphically in the figure below. Equilibrium virulence occurs where a line, rooted at the origin, is tangent to the curve relating transmissibility to disease-induced mortality. When the curve is shifted down, the equilibrium mortality increases. EVD is a zoonosis and it's reasonable to think that when it makes the episodic jump into human populations, it is leaving the reservoir species the biology of which it is adapted to and entering a novel species to which it is not adapted. Transmission efficiency very plausibly would decrease in such a case and we would expect higher virulence.
The second generality that may be of interest for EVD is discussed by Paul Ewald in his book on the evolution of infectious disease and (1998) paper. Ewald notes that when pathogens are released of the constraint between transmissibility and mortality -- that is, when being really sick (or even dead) does not necessarily detract from transmission of the pathogen -- then virulence can increase largely without bound. Ewald uses the difference in virulence between waterborne and directly-transmitted pathogens to demonstrate this effect. At first glance, this seems to contradict the van Baalen & Sabelis model, but it doesn't really. The constraint is represented by the curve in the above figure. When that constraint is released, the downward-sloping curve becomes a straight line (or maybe even an upward-sloping curve) and transmissibility continues to increase with mortality. There is no intermediate optimum, as predicted by the MVT, so virulence increases to the point where host mortality is very high.
A hemorrhagic fever, EVD is highly transmissible in the secretions (i.e., blood, vomit, stool) of infected people. Because these fluids can be voluminous and because so many of the cases in any EVD outbreak are healthcare workers, family members, and attendants to the ill, we might imagine that the constraints between transmissibility and disease-induced mortality on the Ebola Virus could be released, at least early in an outbreak. As behavior changes over the course of an outbreak -- both because of public health interventions and other autochthonous adaptations to the disease conditions -- these constraints become reinforced and selection for high-virulence strains is reduced.
These are some theoretically-informed speculations about the relevance of selection on virulence in the context of EVD. The reality is that while the theoretical models are often supported by experimental evidence, the devil is always in the details, as noted by Ebert & Bull (2003). One thing is certain, however. We will not make progress in our understanding of this horrifying and rapidly changing epidemic if all we are worried about is the virus mutating.
Selection is overwhelmingly the most powerful force shaping evolution. The selective regimes that pathogens face are affected by the physical and biotic environments in which pathogens are embedded. Critically, they are also shaped by host behavior. In the case of the current West African epidemic of EVD, the host behavior in question is that of many millions of people at risk, their governments, aid organizations, and the global community. People have a enormous potential to shape the selective regime that will, in turn, shape the pathogen that will infect future victims. This is what we need to be worrying about, not whether the virus will mutate. It saddens and frustrates me that we live in a country where evolution is so profoundly misunderstood that even our most esteemed, and otherwise outstanding sources of information and opinion don't understand the way nature works and the way that human agency can change its workings for our benefit or detriment.
It's been a long time since I've written in monkey's uncle. Life has gotten pretty busy and my seeming inability to write brief entries has led me to neglect the blog this year. However, I am freshly back from the Ecology and Evolution of Infectious Disease Conference in Fort Collins, Colorado and feel compelled to give my annual run-down. The conference was hosted by friend and colleague Mike Antolin, Sue Vandewoude, and my erstwhile post-doc, now CSU researcher, Dan Salkeld. Nice job, folks, on a very successful conference.
EEID is pretty much the best meeting. As I noted in last year's post, I love its future-orientation. EEID is a meeting that foregrounds the work of junior scientists and there was, as ever, a tremendous array of human capital on display at this meeting. This drives home to me the importance of investment in professional training and research programs that specifically develop human capital. This community exists in large measure because of the innovative program jointly offered by NSF and NIH. Thanks as ever to the vision and hard work of Josh Rosenthal, Sam Scheiner, and all the funders (e.g., support from The Gates Foundation can be found all around this conference) for this area. It's always great to catch up with smart, fun friends. Plenty of time was spent talking science and drinking craft beer (what a beer town Ft. Collins is!) with the likes of Peter Hudson, Jessica Metcalf, Ottar Bjornstad, Aaron King, Mike Antolin, Tony Goldberg, Issa Cattadori, Maciej Boni, Marm Kilpatrick and, of course, Dan Salkeld. It was nice to meet and chat, if only briefly, with my sometime remote collaborator Paul Sharp, who gave what I understand to be an extremely stimulating keynote on the complicated and surprising evolution of malaria (alas, I missed it as I was delayed getting to Ft. Collins). I also spent some quality time learning about acquired immunity in dogs with Colin Parrish. This may come in handy for some ideas that Jess Metcalf and I have been playing around with.
There is a great tradition of the EEID hike and closing banquet/dance. Ft. Collins provided a beautiful and challenging hike out in Lory State Park. The view from the top of Arthur's Peak was pretty amazing.
View from the top of the trail on Arthur's Peak, Lory State Park, Ft. Collins.
At Wednesday's banquet, I'm afraid to say that Princeton once again dominated the dance floor as we all rocked out to the amazing Denver funk/rock/jam band Kinetix (great choice, Mike). The Stanford showing was disappointing in part because of the early departure of some of our most enthusiastic dancers. Don't get cocky though, Princeton. We'll be gunning for you next year.
The entirety of Tuesday morning's session was given over to communicating science. Dan Salkeld warmed up the crowd with some hilarious examples of the reporting frenzy that ensued following the publication of our paper on plague dynamics in prairie dog towns or, more recently, Hillary Young's work showing that excluding large ruminants increases rodent density in Kenya. Wow. Dan also used my Stanford colleagueRebecca Bird's work as an example of how an unexpected story can engage readers and listeners. My collaborator Tony Goldberg gave a talk that was also not lacking in ridiculous headlines thanks to his "viral" nose-tickwork. David Quammen, author of outstanding popular science books such as The Song of the Dodo and Spillover (which Bill Durham and I use for our class on environmental change and emerging infectious disease), gave a terrific presentation in which he consolidated a lot of nice, practical advice on the craft of writing engaging work into 18 points, amply illustrated by anecdotes of characters from our field. Sonia Altizer from the University of Georgia introduced the crowd to the opportunities (and pitfalls) of citizen science and suggested that it might just be possible to engage the public in disease ecology data collection. Some examples she identified included the granddaddy of citizen-science in the US run by the Laboratory of Ornithology at Cornell, the ZomBee Watch at SFSU, and her own Project MonarchHealth. If I had to summarize this session in one pithy phrase, I think it would have to be "Yay, ecologists!"
Quammen took to Twitter to call us out for being behind the curve with respect to social media.
@lfoquet I'll ask. But I think #EEID2014 is a quietish conference, not high on podcasts or social media. Tell me, others, if I'm wrong.
While there were, in fact, a few of us tweeting the occasional tidbit from the conference, I think his general point is valid. This stuff is intrinsically interesting and we can do a much better job communicating to broad publics.
Some talks that really caught my attention.
Ary Hoffmann gave a great talk about the complexities of using bacteria of the genus Wolbachia to control the Aedes mosquitoes that transmit dengue in Australia (and elsewhere). Wolbachia infects mosquitoes and can have a variety of effects on their biology. The reason artificial infection of mosquitoes wit this bacterium seems so promising as a means of biological control is that the offspring of crosses between infected and uninfected mosquitoes are not viable. This is obviously a very substantial fitness cost to the mosquitoes and this creates serious challenges for getting the infected mosquitoes to persist and take over local populations. Hoffmann presented a cool result about the invasibility of infected mosquitoes wherein in the early phases of introduction there is an unstable point in the mosquito dynamics. At this point, if the infected mosquitoes are above a threshold, they will successfully invade, otherwise, they will die out because of the inherent fitness costs of the Wolbachia infection. One policy challenge that arises is that to get a local population of mosquitoes above the invasibility threshold, researchers and vector-control specialists have to sometimes introduce a lot of mosquitoes. This means that the number of mosquitoes locally can increase substantially and, as you can imagine, this isn't always popular with communities.
Fellow Anthropologist Aaron Blackwell from UCSB gave a fantastic talk on our "old friends", the helminths (cue the freaky electron micrograph of a helminth's mouth!). Aaron participates in the Tsimane Health and Life History Project which was started by colleagues Mike Gurven (also at UCSB) and Hilly Kaplan (New Mexico). Using sophisticated multi-state Markov hazard models (go Anthropology!), Aaron showed that co-infection with helminths and Giardia is less frequent than expected among this population that experiences ubiquitous exposure to both pathogens and that, in fact, infection with the one appears to be protective against infection with the other. One of the most provocative results he presented showed that helminth infection actually lowered systolic blood pressure in men by an amount equivalent to the increase that comes from aging ten years. Chronic helminthic infection may be a reason why Tsimane men's systolic blood pressure does not rise precipitously with age as it does in the US. This result, which may provide fresh insights into the mechanisms of hypertension, a major source of morbidity in the US, struck me as particularly poignant given the demeaning comments made about NSF funding for work among the Tsimane from none other than Lamar Smith (R–TX), the chair of the House Committee on Science, Space, and Technology.
Anna Savage, a post-doc with the National Zoo in Washington DC, gave an awesome talk on the comparative immunogenetics of of frogs with respect to infection with the devastating fungal infection, chytridiomycosis. Chytridiomycosis has been identified as a major cause of amphibian extinction worldwide and Anna showed surprising heterogeneity in immune response across frog species. This is a subject with which I have only passing familiarity, but her talk demonstrated an amazing sophistication in integrating different levels of biological organization and making sense of a dauntingly complex problem. I would wager that Dr. Savage is one to keep an eye on.
The organizers tried a scheduling format that was a bit different from last year, wherein each session started with two half-hour talks generally given by somewhat more senior people. The second half of each session was then given over to brief ten-minute talks, typically delivered by more junior people. This format is nicely in keeping with the great EEID tradition of promoting the research of junior scientists. A few short talks that I found especially interesting included one by Sarah Hamer, from Texas A&M, on Chagas disease in the United States. She presented sobering data from national blood-bank surveillance showing a surprising number of Chagas-infected samples coming from donors with no history of travel to Latin America. When pushed by a questioner, she suggested that she would consider Chagas to be endemic in the US, at least in dogs and possibly even in people. Carrie Cizauskas, formerly of Wayne Getz's shop at Berkeley and now with Andy Dobson and Andrea Graham at Princeton, give a nice talk on the role of both stress and sex hormones in mediating macroparasite infection in wild ungulates in Etosha National Park, Namibia. Romain Garnier from Princeton described a very nifty image-processing approach to scanning large volumes of histological slides for indications of infection.
I perhaps didn't see as many posters as I should have. The problem with the poster sessions is that one keeps running into various people one wants to talk to. I did manage to check out the poster of my former freshman advisee and current Princeton EEB student Cara Brook. She's got an awesome PhD project studying the multi-host ecology of infectious disease in Malagasy fruit bats.
I'm looking forward to next year's meeting at the University of Georgia already. I'm also looking forward to resuscitating the pedagogical workshop that used to be a signature feature of this EEID meeting. More on that later...
I am recently back from the 2013 Ecology and Evolution of Infections Disease Conference at Penn State University. This was quite possibly the best meeting I have ever attended, not even for the science (which was nonetheless impeccable), but for the culture. I place the blame for this awesome culture firmly on the shoulders of the leaders of this field and, in particular, the primary motivating force behind the recent emergence of this field, Penn State's Peter Hudson. Since I had attended the other EEID conference at UGA earlier this Spring (another great conference), I had no intention on attending the Penn State conference this year. Then, one day in late March, Nita Bharti asked me if I was going and mentioned, "You know it's Pete's 60th birthday, right?" Well that sealed it; I really had no choice. I simply had to go if for no other reason than to pay my due respect to this man I admire so greatly. Pete has the most relentless optimism about the future of science and a willingness to make things happen that I have ever encountered and, in this way, has provided me one of my primary role models as a university professor and mentor. He has played a role in developing so many of the brilliant people who make this field so exciting, it's amazing (just a sample that comes immediately to mind: Ottar Bjornstad, Matt Ferrari, Nita Bharti, Marcel Salathé, Isabella Cattadori, Jamie Lloyd-Smith, Shweta Bansal, Jess Metcalf...). Of course, even as I write this, I realize the joint influence of another major player in the field, Bryan Grenfell, formerly of Penn State but now at Princeton, becomes obvious. A great scientist in his own right, Pete is the master facilitator, providing the support (and institutional interference!) that allows young scholars to thrive. He is a talent-spotter extraordinaire.
The talks that made up the bulk of the scientific program were, for the most part, excellent. The average age of the speakers was about 30, maybe just a bit higher. When one attends an academic conference, one typically expects that the major addresses to the collected masses will be by geezers, er, senior scholars in the field. There was a clear play at inversion of the standard model here though. Speakers were clearly chosen because of their trajectories, not their past achievements. That's pretty great. When I went up for tenure at Stanford, I was told that Stanford does not really care about what you have done; it cares about what you will do. Of course, the best information that the university has about your future work is the work you have already done. This conference embodied this spirit by placing the future (and, in many cases, current) leaders of the field in the key speaking roles while some of the biggest names in ecology, population biology, and epidemiology sat happily in the audience (e.g., joining Hudson and Grenfell were Andy Dobson, Andrew Read, Mick Crawley, Charles Godfray, Mike Boots, Mercedes Pascual, Les Real, Matt Thomas, ...)
The tone set by these great mentors carries through to the whole culture of the conference, where senior people attended the poster sessions, sat with students at lunches and dinners, and schmoozed at the plentiful open-bar mixers. For example, on the first full day of the conference, there was an afternoon poster session that started at 4:30 (we had been in back-to-back sessions since 8:30). This session was preceded by an hour-long poster-teaser session in which grad students and post-docs got up and presented 60-second (and, as Andrew Read noted, not one nanosecond more) teasers of their posters. Bear in mind, this session was entirely comprised of students and post-docs. It was striking that essentially every seat in the house was occupied and all the major players were present. The teasers were great – many were very funny, including a haiku apparently written by a triatomine bug and translated to us by Princeton EEB student Jennifer Peterson.
After the teasers, the conference went en masse to the fancy new Millenium Science Complex (it turns out that Pete Hudson has physical capital projects in addition to human capital ones!). There, participants milled about the 150 posters. After spending quite a bit of time doing this – and dutifully getting pictures of all my lab with their posters – I thought to check the time and realized it was nearly 6:30. The poster session had been going for two hours and nearly everyone was still there, including all the luminaries. It helped that there was free beer. I tweeted my amazement at this realization:
That is, in fact, Princeton's Bryan Grenfell moving fast in the middle of the picture, apparently making a bee-line for Michigan's Aaron King. Andrew Read is in the far background, talking to a poster-presenter (he has that posture).
Scientific highlights for me included Caroline Buckee's talk about measuring mobility in the context of malaria transmission in Kenya and Derek Cummings's talk on the Fluscape Project to measure spatial heterogeneity in influenza transmission in China. I am a long-time fan of this project and it's nice to see the great work that has come out of it. These talks were right in my wheelhouse of interest, but there were plenty other cool ones including Britt Koskella's talk on the dynamics of bacteria and phage on tree leaves.
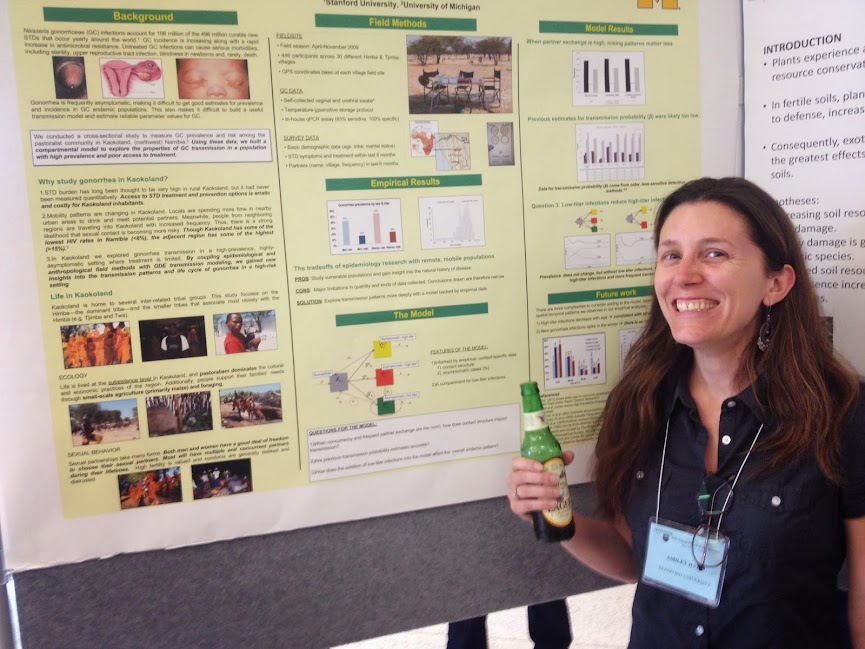
Stanford was exceedingly well represented at this conference. My lab had no fewer than five posters. Ashley Hazel presented on her work with Carl Simon on modeling gonorrhea transmission dynamics in Kaokoland, Namibia. Whitney Bagge presented her work on remote-sensing of rodent-borne disease in Kenya. Alejandro Feged presented work on the transmission dynamics of malaria in the Colombian Amazon among the indigenous Nukak people. Laura Bloomfield presented her remote sensing and spatial analysis work from our project on the spillover of primate retroviruses in Western Uganda. I closed things out with a minimalist poster on simple graphical models for multiple attractors in vector-borne disease dynamics in multi-host ecologies. In addition to my lab group, Giulio De Leo (with whom I have been running a weekly disease ecology workshop at Woods since winter quarter) was there, helping to bridge all sorts of structural holes in our collective collaboration graphs.
The other thing that comes out of these meetings, especially more intimate ones like EEID, is some actual work on collaborative projects. I managed to find some time to sit down and discuss plans with collaborators as well as do some shameless recruitment for my planned re-submission of the Stanford Biodemography Workshops. I'm really excited about some of these collaborations, including one that brings together my two major areas of interest: biodemography and life history theory and infectious disease ecology.
Oh, and I'm convinced that there must be an interpretive dance component to the Ph.D. exam in the Grenfell lab. This is certainly the most parsimonious explanation for much of what I saw Wednesday night.
I'm done now with the first week of the Spring quarter. It was a bit challenging because I had to attend the PAA meetings in Washington, DC for the latter part of the week, but Brian Wood ably covered for me on Thursday. I thought that I would use the blog as a tool for summarizing one of the key points I want students to take away from this fist week in which we discussed evolution and natural selection.
We spent a good deal of lecture time talking about adaptation. Specifically, we discussed how adaptation can serve as a foil to typology and essentialism. Adaptation is local and must be seen within its specific environmental and historical context. Adaptations are dynamic because environments are.
Adaptationist thinking is powerful, but can easily be overdone. This is why I also think it is essential to understand the mechanics of selection, something that I'm afraid is not often addressed in introductory evolutionary anthropology classes. So, in the very first lecture of class, I throw some quantitative genetics (and, thus, some math) at students. Of course, these are Stanford students, so I'm confident they can handle a little techie-ness every now and then. We specifically discuss the multivariate breeder's equation, sometimes known as Lande's equation:
,
where is the change in the mean fitness of a multivariate trait, is the additive genetic variance-covariance matrix, and is the selection gradient on .
In effect, is a vector pointing in the direction of the optimal change in the phenotype. The matrix does two things to this gradient pushing toward its optimum: (1) it scales the response depending on how much additive variance there is in each trait and (2) it rotates it as a function of the covariances between traits. I won't get too much into matrix multiplication here (this is a very nice reference too). The key point is that is a square matrix (where is the number of traits we're looking at) the diagonal elements of which are variances and the off-diagonal elements of which, represent the covariances between traits and . Selection requires variance. Without sufficient variance, even strong selection won't change the phenotype much between generations. But variance isn't all there is to it. When the covariances are positive, there will be substantial indirect selection, and when they are negative, you have genetic constraints at work. Selection may be pointing in a particular direction, but the structure of the trade-offs could very easily mean that you can't actually get there.
Let's consider three quick (toy) examples. Say we have two traits, maybe "length" and "width" (this could be something less vague and insipid: Lande (1979) looks at brain mass and body mass in a serious two-trait example). We will assume that the selection gradient is . That is, the force of selection is twice as high on length as it is on width, but it is pretty strong and positive on both. We'll demonstrate the effect of variance and constraint in three ways: (1) more variance in the trait under weaker selection (), (2) positive covariance between the two traits (), and (3) negative covariance between the two traits ().
The figure below plots the response to selection in the three different types of genetic architecture. The direction of selection is indicated in the grey arrow. If the variances of the two traits were equal to 1 and there were zero covariances, this is where selection would move the phenotype pair (try it). We can see that the response to selection moves toward width (the trait under weaker selection) even when covariances are zero (black arrow). Why? Because there is more variance for width than there is for length (). This effect becomes more pronounced when there is positive covariance between the traits (blue arrow) -- the selection toward width is . When the covariances are negative, we see something cool (red arrow). The response to selection is small and moves (almost) entirely in the direction of length. This is because the negative covariance between length and width, when acted on by the strong selection on length, all but cancels out the positive response to selection ().
This simple demonstration shows that the response to selection can be complex. Making an argument that some trait would be under selection is not sufficient to say that it actually evolved (or will evolve) that way. Entirely plausible arguments for the direction of selection are made all the time in evolutionary anthropology. Here is one from a very important paper in paleoanthropology (Lovejoy 1981: 344):
Any behavioral change that increases reproductive rate, survivorship, or both, is under selection of maximum intensity. Higher primates rely on social behavioral mechanisms to promote survivorship during all phases of the life cycle, and one could cite numerous methods by which it theoretically could be increased. Avoidance of dietary toxins, use of more reliable food sources, and increased competence in arboreal locomotion are obvious examples. Yet these are among the many that have remained under stadong selection throughout much of the course of primate evolution, and therefore unlikely that early hominid adaptation was a product of intensified selection for adaptations almost universal to anthropoid primates.
Arguing for selection without considering trade-offs can get you into trouble. Selection in the presence of quantitative genetic constraints (or even differential variance in the traits) can produce counter-intuitive results. (Selectionists, don't dispair. There are ways to deal with this, but it will have to wait for another post). In the case of Lovejoy's argument, there are good reasons to think that survivorship and reproductive rate are, indeed, strongly negatively correlated. Which is under stronger selection? Which has more additive variance? How strong are the negative covariances?
When we make selectionist or adaptationist arguments, we should always keep in the back of our minds the three questions:
How strong is the force of selection?
How much variance is there on which selection can act?
How is the trait constrained through negative correlations with other traits?
References
Lande, R. A. 1979. Quantitative genetic analysis of multivariate evolution applied to brain: body size evolution. Evolution. 33:402-416.
Lovejoy, C. O. 1981. The origin of man. Science. 211:341-350.
Nicholas Wade, who normally writes really terrific stuff on science in the New York Times, has a brief piece on our Anthropology fracas du jour. It's good to see an expression of concern for the place of science in anthropology in such a prominent place and by such an important science writer. I just wish he had gotten a few more things right. While the Darkness in El Dorado fiasco was not a high point for the AAA, I suspect that this had not one iota to do with the re-wording of AAA's long-range planning document. Secondly, I was pretty horrified to learn that science can't be used as a framework for studying gender, ethnicity, and race, nor, apparently, can scientists advocate for indigenous people's or human rights:
The decision [to remove the word 'science' from the long-range planning document] has reopened a long-simmering tension between researchers in science-based anthropological disciplines — including archaeologists, physical anthropologists and some cultural anthropologists — and members of the profession who study race, ethnicity and gender and see themselves as advocates for native peoples or human rights.
I think that this will come as quite an unpleasant surprise to many fine scientific anthropologists who are apparently fooling themselves by attempting to understand race or gender or working to improve the lives of the people with whom they work.
So, I'm left with mixed feelings about this turn of events. On the one hand, the prominence of a Science Times piece by Nicholas Wade means that debate is likely to continue for a while to come. It would be particularly helpful if this work helped engage what I suspect is a quiet majority of anthropologists who are (1) sympathetic to science maintaining a prominent place in anthropology, and (2) too busy with their work to worry about yet another shrill controversy in the professional society they may or may not belong to (having given up membership because they already felt it didn't represent their interests). On the other hand, I think we're going to need to stop being inflammatory and falling back on facile received categories (e.g., "postmodernists," "sociobiologists," etc.) at every opportunity if we are going to make this debate productive and fashion a society that is friendly to rigorous scholarship in whatever form it may take. For my part, I am sticking with my view that the best way to promote science in anthropology is to do it, do it well, and communicate with a broad scientific readership.
One of the fundamental ontological questions of our day is surely, "is there anything you can't do with ape scat?" Well, OK, this might be pushing it a bit far, but a recent article in the New York Times makes a pretty strong case for the blessings of this pungent goo. My collaborator Beatrice Hahn, quoted in this article as saying that ape scat is "worth its weight in gold," has been collecting fecal samples gathered by far-flung ape researchers throughout Africa. In addition to providing fundamental data on the landscape-level distribution of SIV (the work on which I have collaborated with her), Beatrice's ape scat collection has now yielded the secret of the origin of Plasmodium falciparum, the most deadly of the five species that cause human malaria infection. The paper by Liu and colleagues appeared in the 23 September issue of Nature. There is a nice accompanying piece by Eddie Holmes as well. It turns out that P. falciparum malaria spilled over into human populations from western gorillas, rather than from chimpanzees as had long been thought. Makes all that smelly collecting actually seem worthwhile...
My peeve is the use of the Latin binomial for our species in the caption of the figure comparing an anatomically modern human skeleton with a reconstruction of a Neanderthal skeleton. The caption reads:
A reproduction of a Neanderthal skeleton, left, and the original modern homo sapien skeleton, right.
Egads! First, our genus is, of course, Homo, not homo (note the capitalization and some sort of typographic elaboration, either italics or underline to denote the special status of a Latin binomial). Second, the species name is sapiens not sapien.
Systematic nomenclature is abused all the time. I shudder every time I read the ingredients list on the Aveda shampoo in our shower (ingredient #3 of the clove shampoo is written "Prunus Amydalus Dulcis (Sweet Almond)"). It's nice that they are trying to be specific and precise about the composition of their product, but get the naming conventions right! Even if it's impractical to italicize the text for printing reasons, please remember, genera are capitalized; species (and sub-species) are lower case.
Why am I so uptight about such a seemingly trivial issue of typography and convention? It's because nomenclature matters in science. In particular, I think that systematic nomenclature (i.e., nomenclature that describes evolutionary relationships) does for biology what Bertrand Russell argued good formal notation does for mathematics and logic: provides a subtlety and suggestiveness that allows it to almost teach for itself.
Matt Ridley pens a hilarious simile in his great book, Nature Via Nurture(published as The Agile Gene in the United States) that I think you might actually need to be an evolutionary anthropologist to fully appreciate. And I quote:
Just as sex enabled mammals to combine two great inventions -- lactation and the placenta -- so trade enabled early people to combine draft animals and wheels to better effect. (Ridley 2003: 228)
Just like it. Awesome!
notes on human ecology, population, and infectious disease



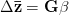
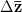
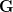
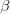
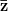
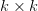
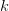
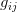
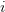
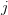

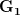
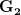
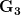
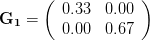
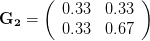
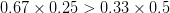
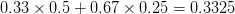
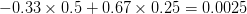

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}