OK, not exactly new new, but certainly newish. This post is part of my new attempt to update my blog more with stories about science, research, and education in an attempt to avoid the vicious cycle of anxiety and depression that comes from spending too much time reading the news and engaging in social media.
Our paper on the prevalence of asymptomatic Ebola cases in Sierra Leone was published in PLoS Neglected Tropical Diseases on 15 November. In it, we show that nearly 10% of a sample of people from an Ebola hotspot in Kono District, Sierra Leone, tested positive for Ebola virus antibodies despite having reported no symptoms of Ebola Virus Disease.
The West African Ebola epidemic of 2014-2015 was the biggest outbreak of the disease ever recorded, with over 28,000 reported cases. Our results suggest that the total number of cases may have been quite a bit more than this. They also suggest that Ebola is, as we suspected, like other pathogens and causes a wide variety of clinical manifestations.
This work was led by my rock-star Ph.D. student, Gene Richardson and involved a great many collaborators. It was a great honor to be able to publish with such luminaries as George Rutherford, Megan Murray, and Paul Farmer. With several papers in the works or already submitted and ongoing research, I'm really looking forward to more results in the near future!
About this time of the year, I generally do a re-cap of the American Anthropological Association's annual meeting. However, I didn't attend AAAs this year for the first time in five years, so I don't have much to report. Anthropologists' annual awkwardly-timed professional ritual just went down in Washington DC and I thought I would see if anything newsworthy came of it. Doing a Google news search with a variety of permutations of the association name (American Anthropological Association in quotes and not, AAA, etc.) and other keywords (Washington, annual, meeting, 2014, etc.), I managed to find one or two things. As I (and others) have noted before, the AAA meetings don't attract a lot of press. New discoveries or items of broad public interest are apparently not generally discussed at AAA. This year, the most notable item in a news search is the rejection of a resolution to boycott Israel over what the resolution referred to as “Israel’s ongoing, systematic, and widespread violations of Palestinian academic freedom and human rights.”
One other item popped up which actually resembles something newsworthy on the scholarly front (as opposed to the business of the association). Kari Lyderson at the Crux writes about a movement to bring anthropological expertise to bear on the ongoing Ebola Virus Disease epidemic in West Africa. Sharon Abramowitz, a terrific medical anthropologist at the University of Florida, has helped to found an initiative called the Ebola Emergency Response Initiative, the aim of which is to provide social and cultural expertise to help with control of the EVD epidemic. This is good news and exactly the sort of thing I would like to see more of at AAA. There are many ways that improved cultural understanding by medical personnel and public health practitioners could help to bring this epidemic under control – a point that anthropologist/human behavioral ecologist Barry Hewlett been making for years now. These are issues we've thought about a bit here and that my Ph.D. student Gene Richardson is actively working on in Sierra Leone right now.
I am just back from an event at the University of California, Irvine organized by medical demographer Andrew Noymer. The event drew a big crowd, with probably 500-600 people in attendance.
There were five invited plenary speakers: Michael Buchmeier (UCI) spoke about the virology of Ebola and the Filovriuses more generally. Hearing Mike's insights on the not one, but two vaccines for Ebola that have been shelved for a decade due to lack of interest was particularly illuminating. George Rutherford (UCSF) talked about the epidemiology of the current EVD epidemic and placed control efforts within the broader context of Global Health Initiatives. This is a guy with a ton of experience in global health and on the ground in Africa and his cool demeanor was calming for the crowd. Victoria Fan (Hawai'i) discussed the economic implications of the epidemic. Spoiler alert: they're not good. Shruti Gohil (UCI Medical Center) talked about infection control in a hospital setting. Finally, I talked about the disease ecology, broadly construed, of Ebola. Following our talks, we got together as a panel and took questions for the audience.
Given the crazy hysteria surrounding the EVD epidemic and the arrival of a handful of cases in the United States, it was reassuring to participate in a couple hours of such sober, scientifically-informed discussion. Shruti's insights as chief of infection control at the UCI medical center particularly struck me. She noted that Texas Health Presbyterian Hospital in Dallas, where the first American EVD case (Thomas Duncan) was treated, was clearly completely unprepared to handle an acute EVD case. Despite this, Shruti estimated that the attack rate of health care workers who attended to Duncan was about 4%. Not that horrible for an unprepared hospital. She also noted that no health care workers have become infected in the special units specifically designed to handle infectious diseases like EVD at Emory, Nebraska, and Bethesda. Planning, strict adherence to protocols, and personal protective gear work!
So, let's summarize a bit about EVD in the US (these are the numbers as best as I can remember them, with citations where I can find them):
Another interesting point that Shruti made is that none of Duncan's close personal contacts have contracted EVD and the 21-day window has now passed. The clear implication of all these data is that Ebola is not that infectious. It is controllable if we are prepared and follow protocols.
This gives me hope that we can control the EVD epidemic in West Africa if we were to decide to get serious about its control. But the international community needs to fight this epidemic where it is currently raging. This is clearly in the national interest of the United States and the collective interest of the international community. If we want to remain secure from EVD, we need to stop it where the epidemic continues to grow. World Bank President, and medical anthropologist extraordinaire, Jim Kim pulled out a great analogy in an interview on NPR on 17 October:
It's like you're in your room and the house is on fire, and your approach is to put wet towels under the door. That might work for a while, but unless you put the fire out, you're still in trouble.
Let's get over our fear, stop politicizing this crisis, stop demonizing the heroes. Let's roll up our sleeves, get out our checkbooks, and bring a speedy end to this crisis. Let's put out the fire.
I just read an excellent piece at Slate.com this morning by Benjamin Hale. He notes that the scariest, most insidious thing about Ebola Virus Disease is that the disease capitalizes on intimate contact for transmission. While diseases such as influenza or cholera are transmitted by casual contact, frequently to strangers, via aerosolized droplets (influenza) or fecally contaminated water (cholera). Caretakers, and especially women, are hit hard by EVD. Hale writes,
...the mechanism Ebola exploits is far more insidious. This virus preys on care and love, piggybacking on the deepest, most distinctively human virtues. Affected parties are almost all medical professionals and family members, snared by Ebola while in the business of caring for their fellow humans. More strikingly, 75 percent of Ebola victims are women, people who do much of the care work throughout Africa and the rest of the world. In short, Ebola parasitizes our humanity.
True, and tragic, enough. But this article falls prey to one of my biggest frustrations with the reporting of science, one that I have written about recently in the context of the current EVD epidemic ravaging West Africa.
In the list Hale presents of the major concerns about EVD, he notes: "The threat of mutation," citing concern that Ebola virus might become airborne in a news report in Nature and the New York Times article that got me so worked up 10 days ago. Earlier this week, there was yet another longish piece in Nature/Scientific American that mentions "mutation" seven times but never once mentions selection. Or in another Nature piece, UCSF infectious disease physician Charles Chiu is quoted: "The longer we allow the outbreak to continue, the greater the opportunity the virus has to mutate, and it’s possible that it will mutate into a form that would be an even greater threat than it is right now.” True, mutations accumulate over time. Not true, mutation alone will make Ebola virus a greater threat than it is now. That would require selection.
While the idea of airborne transmission of Ebola virus is terrifying, the development of the ability to be transmitted via droplet or aerosol would be an adaptation on the part of the virus. Adaptations arise from the action of selection on the phenotypic variation. Phenotypes with higher fitness come to dominate the population of entities of which they are a part. In the case of a virus such as Ebola virus, this means that the virus must make sufficient copies of itself to ensure transmission to new susceptible hosts before killing the current host or being cleared by the host's immune system. While efficient transmission of EVD by aerosol or droplet would be horrible, equally horrible would be an adaptation that allowed it to transmit more efficiently from a dead host. It's not entirely clear how long Ebola virus can persist in its infectious state in the environment. In a study designed to maximize its persistence (indoors, in the dark, under laboratory conditions), Sagripanti and colleagues found that Ebola virus can persist for six days. Under field conditions, it's probably much shorter, but CDC suggests that 24 hours in a reasonably conservative estimate.
The lack of a strong relationship between host survival and pathogen transmission is why cholera can be so devastatingly pathogenic. The cholera patient can produce 10-20 liters of diarrhea (known as "rice water stools") per day. These stools contain billions of Vibrio cholerae bacteria, which enter the water supply and can infect other people at a distance well after the original host has died. The breaking of the trade-off between host mortality and the transmissibility of the pathogen means that the natural break on virulence is removed and the case fatality ratio can exceed 50%. That's high, kind of like the current round of EVD. Imagine if the trade-off between mortality and transmission in EVD were completely broken...
Changes in pathogen life histories like increased (or decreased) virulence or mode of transmission arise because of selection, not mutation, and this selection results from interactions with an environment that we are actively shaping. Sure, mutation matters because it provides raw material upon which selection can act, but the fact remains that we are talking primarily about selection here. Is this pervasive misunderstanding of the mechanisms of life the result of the war of misinformation being waged on science education in the US? I can't help but think it must at least be a contributor, but if it's true, it's pretty depressing because this misunderstanding is finding its way to some of the world's top news and opinion outlets.
This has to be a quick one, but I wanted to go on the record is noting my frustration at the current concern that Ebola might "mutate" into something far worse, like a pathogen that is efficiently transmitted by aerosol. For example, Michael Osterholm wrote in the New York Times yesterday, "The second possibility is one that virologists are loath to discuss openly but are definitely considering in private: that an Ebola virus could mutate to become transmissible through the air." I heard Morning Edition host David Greene ask WHO Director Margaret Chan last week, "Is this virus mutating in a way that could be very dangerous, that could make it spread faster?"
I agree, Ebola Virus becoming more easily transmitted by casual contact would be a 'nightmare scenario.' However, what we need to worry about is not mutation per se, but selection! Yes, the virus is mutating. It's a thing that viruses do. Ebola Virus is a Filovirus. It is composed of a single strand of negative-sense RNA. Like other viruses, and particularly RNA viruses, it is prone to high mutation rates. This is exacerbated by the fact that RNA polymerases lack the ability to correct mistakes. So mutations happen fast and they don't get cleaned up. Viruses also have very short generation times and can produce prodigious copies of themselves. This means that there is lots of raw material on which selection can act, because variation is the foundation of selection. Add to that heritability, which pretty much goes without saying since we are talking about the raw material of genetic information here, and differential transmission success and voilà, selection!
And virulence certainly responds to selection. There is a large literature on experimental evolution of virulence. See for example the many citations at the linked to Ebert's (1998) review in Sciencehere. There are lots of different specific factors that can favor the evolution of greater or lesser virulence and this is where theoretical biology can come in and make some sense of things. Steve Frank wrote a terrific review paper in 1996, available on his website, that describes many different models for the evolution of virulence. Two interesting regularities in the the evolution of virulence may be relevant to the current outbreak of EVD in West Africa. The first comes from a model developed by van Baalen & Sabelis (1995). Noting that there is an inherent trade-off between transmissibility of a pathogen and the extent of disease-induced mortality that it causes (a virus that makes more copies of itself is more likely to be transmitted but more viral copies means the host is sicker and might die), they demonstrate that when the relative transmissibility of a pathogen declines, its virulence will increase. They present a marginal value theorem solution for optimal virulence, which we can represent graphically in the figure below. Equilibrium virulence occurs where a line, rooted at the origin, is tangent to the curve relating transmissibility to disease-induced mortality. When the curve is shifted down, the equilibrium mortality increases. EVD is a zoonosis and it's reasonable to think that when it makes the episodic jump into human populations, it is leaving the reservoir species the biology of which it is adapted to and entering a novel species to which it is not adapted. Transmission efficiency very plausibly would decrease in such a case and we would expect higher virulence.
The second generality that may be of interest for EVD is discussed by Paul Ewald in his book on the evolution of infectious disease and (1998) paper. Ewald notes that when pathogens are released of the constraint between transmissibility and mortality -- that is, when being really sick (or even dead) does not necessarily detract from transmission of the pathogen -- then virulence can increase largely without bound. Ewald uses the difference in virulence between waterborne and directly-transmitted pathogens to demonstrate this effect. At first glance, this seems to contradict the van Baalen & Sabelis model, but it doesn't really. The constraint is represented by the curve in the above figure. When that constraint is released, the downward-sloping curve becomes a straight line (or maybe even an upward-sloping curve) and transmissibility continues to increase with mortality. There is no intermediate optimum, as predicted by the MVT, so virulence increases to the point where host mortality is very high.
A hemorrhagic fever, EVD is highly transmissible in the secretions (i.e., blood, vomit, stool) of infected people. Because these fluids can be voluminous and because so many of the cases in any EVD outbreak are healthcare workers, family members, and attendants to the ill, we might imagine that the constraints between transmissibility and disease-induced mortality on the Ebola Virus could be released, at least early in an outbreak. As behavior changes over the course of an outbreak -- both because of public health interventions and other autochthonous adaptations to the disease conditions -- these constraints become reinforced and selection for high-virulence strains is reduced.
These are some theoretically-informed speculations about the relevance of selection on virulence in the context of EVD. The reality is that while the theoretical models are often supported by experimental evidence, the devil is always in the details, as noted by Ebert & Bull (2003). One thing is certain, however. We will not make progress in our understanding of this horrifying and rapidly changing epidemic if all we are worried about is the virus mutating.
Selection is overwhelmingly the most powerful force shaping evolution. The selective regimes that pathogens face are affected by the physical and biotic environments in which pathogens are embedded. Critically, they are also shaped by host behavior. In the case of the current West African epidemic of EVD, the host behavior in question is that of many millions of people at risk, their governments, aid organizations, and the global community. People have a enormous potential to shape the selective regime that will, in turn, shape the pathogen that will infect future victims. This is what we need to be worrying about, not whether the virus will mutate. It saddens and frustrates me that we live in a country where evolution is so profoundly misunderstood that even our most esteemed, and otherwise outstanding sources of information and opinion don't understand the way nature works and the way that human agency can change its workings for our benefit or detriment.
I've been traveling all summer while this largest Ebola Virus Disease (EVD) outbreak in recorded history has raged in the West African countries of Guinea, Sierra Leone, Liberia, and (worryingly) Nigeria. My peripatetic state has meant that I haven't been able to devote as much attention to this outbreak as I would like to. There is a great deal of concern -- some might say hysteria -- about EVD and the possibility that it may go pandemic. Tara Smith at least, on her Aetiology blog, has written something sensible, noting that EVD, while terrifying, is controllable with careful public health protective measures, as the historical record from Uganda shows. A recent post by Greg Laden got me to thinking about the numbers from the current EVD outbreak and what we might be able to learn.
EVD was the model disease for the terrible (1995) Dustin Hoffman movie, Outbreak. As we learned in the much more scientifically-accurate (2011) movie Contagion (which is based on an equally terrifying aerosolized Nipah virus), one of the key pieces of information regarding an epidemic is the basic reproduction number, . The basic reproduction number tells us how many secondary infections are expected (i.e., on average) to be produced by a single, typical case at the outset of an epidemic before the pool of susceptible people has been depleted. provides lots of information about epidemics, including: (1) the epidemic threshold (i.e., whether or not an epidemic will occur, which happens in the deterministic case when ), (2) the initial rate of increase of an epidemic, (3) the critical vaccination threshold (i.e., what fraction of the population you need to vaccinate to prevent an outbreak), (4) the endemic equilibrium of an infection (i.e., the fraction of the population that is infected in between outbreaks), and (5) the final size of the epidemic (i.e., the fraction of the total population that is ever infected when the epidemic is over).
Thus, for a novel outbreak, it's good to have an idea of . I've been a bit out of the loop this summer and haven't seen any estimates so I figured that I would see what I could do. I fully realize that someone may have already done this and that I am not yet aware of it. I also recognize that, if someone has done this, they've probably done it better. This is a blog, not a peer-reviewed paper, and I am away from my usual resources, so please take this in the back-of-the-envelope spirit in which it is intended. I reserve the right to retract, etc. I will also post the R code that I used to make the calculations. I hope that this may prove helpful to others interested in the dynamics of outbreaks.
In their terrific (2003) paper on the SARS outbreak, Marc Lipsitch and colleagues provided a method for estimating the reproduction number from outbreak data. Note that this is a more generalized reproduction number, which we call , than is the basic reproduction number, . The key difference is that a reproduction number can be calculated at any point in an outbreak, whereas is only technically correct at the outset (the zero index in indicates the "generation" of the outbreak where "0" refers to the index case, a.k.a., "patient zero"). I've simply used the count of total cases from this week. It is straightforward to extend the calculation to previous counts. I haven't yet had a chance to do this because there is no convenient collection of data that I can find with my current access constraints.
The method involves equating for a simplified SEIR system to the observed rate of increase of the outbreak at some point in time , using the fact that the reproduction number is approximately equivalent to the growth rate of the epidemic. See the supplementary information from Lipsitch et al. (2003) for details of the method. In brief, we calculate the dominant eigenvalue of the linearized SEIR model, for which it is straightforward to write an analytical formula, and equate this to , the empirical growth rate of the epidemic (where is the cumulative number of cases at time ). Lipsitch et al. (2003) note that using the standard formula for the characteristic equation of the eigenvalues of the linearized SEIR model, we can solve for the reproduction number as:
where is the serial interval (i.e., the summed duration of the incubation period, , and the duration of the infectious period, ), is the positive root of the characteristic equation which we set equal to , and is the ratio of the infectious period of the serial interval.
I got the case data from the weekly WHO outbreak report for 11 August 2014. For this week . For the start time of the epidemic in the currently afflicted countries, I used the date of 10 March 2014, taken from this week's NEJM paper by Blaize et al. (2014). For the serial interval data, I used the values provided by the Legrand et al. (2007). Because Legrand et al. (2007) provide mean values of the relevant parameters -- and this is a different epidemic -- I used a variety of values for and to calculate . It turns out that it doesn't matter all that much; the estimates of are pretty stable.
I plot the values of against the duration of the latent period. The different lines are for the different values of the duration of infectiousness. increases with both. What we see is that at this point in the epidemic at least, ranges from around 1.3 to 2.6, depending on specifics of the course of the disease. This is not all that high -- about the same as various flavors of influenza and considerably less than, say, pertussis. This is good news for potential control, if we could just rally some more international support for control of this serious infection...
Here is the R code for doing the calculations and creating this figure:
[r]
library(lubridate)
# number of cases as of 11 August 2014
# http://www.who.int/csr/don/2014_08_11_ebola/en/
cases <- 1848
# start of epidemic in Guinea: 10 March 2014
# Blaize et al. (2014), NEJM. DOI: 10.1056/NEJMoa1404505
s <- dmy("10-03-14")
e <- dmy("11-08-14")
t <- e-s
# Time difference of 154 days
## incubation period 2-21 days
## http://www.who.int/mediacentre/factsheets/fs103/en/
## duration of infectiousness: virus detected in of lab-infected man 61 days!
## Legrande et al. (2007) use L=7 and D=10
## doi:10.1017/S0950268806007217
lambda <- log(cases)/t
## From Lipsitch et al. (2003)
## lambda is the dominant eigenvalue of the linearized SEIR model
## V is the serial interval V = D + L
## D is duration infectious period, L is duration of latent period
## f is the ratio of the the infectious period to the serial interval
## to solve for R set the eigenvalue equal to the observed exponential growth rate of the epidemic log(Y(t))/t
Rapprox <- function(lambda,V,f) 1 + V*lambda + f*(1-f)*(V* lambda)^2
RR <- matrix(0, nr=10, nc=10)
L <- seq(3,12)
D <- seq(5,14)
for(i in 1:length(L)){
for(j in 1:length(D)){
RR[i,j] <- Rapprox(lambda,L[i]+D[j],D[j]/(L[i]+D[j]))
}
}
cols <- topo.colors(10)
png(file="Ebola-R0-plot1.png")
plot(L, RR[1,], type="n", xlab="Duration of Incubation", ylab="Reproduction Number",ylim=c(1,2.5))
for(i in 1:10) lines(L, RR[i,], lwd=2, col=cols[i])
dev.off()
[/r]
It's been a long time since I've written in monkey's uncle. Life has gotten pretty busy and my seeming inability to write brief entries has led me to neglect the blog this year. However, I am freshly back from the Ecology and Evolution of Infectious Disease Conference in Fort Collins, Colorado and feel compelled to give my annual run-down. The conference was hosted by friend and colleague Mike Antolin, Sue Vandewoude, and my erstwhile post-doc, now CSU researcher, Dan Salkeld. Nice job, folks, on a very successful conference.
EEID is pretty much the best meeting. As I noted in last year's post, I love its future-orientation. EEID is a meeting that foregrounds the work of junior scientists and there was, as ever, a tremendous array of human capital on display at this meeting. This drives home to me the importance of investment in professional training and research programs that specifically develop human capital. This community exists in large measure because of the innovative program jointly offered by NSF and NIH. Thanks as ever to the vision and hard work of Josh Rosenthal, Sam Scheiner, and all the funders (e.g., support from The Gates Foundation can be found all around this conference) for this area. It's always great to catch up with smart, fun friends. Plenty of time was spent talking science and drinking craft beer (what a beer town Ft. Collins is!) with the likes of Peter Hudson, Jessica Metcalf, Ottar Bjornstad, Aaron King, Mike Antolin, Tony Goldberg, Issa Cattadori, Maciej Boni, Marm Kilpatrick and, of course, Dan Salkeld. It was nice to meet and chat, if only briefly, with my sometime remote collaborator Paul Sharp, who gave what I understand to be an extremely stimulating keynote on the complicated and surprising evolution of malaria (alas, I missed it as I was delayed getting to Ft. Collins). I also spent some quality time learning about acquired immunity in dogs with Colin Parrish. This may come in handy for some ideas that Jess Metcalf and I have been playing around with.
There is a great tradition of the EEID hike and closing banquet/dance. Ft. Collins provided a beautiful and challenging hike out in Lory State Park. The view from the top of Arthur's Peak was pretty amazing.
View from the top of the trail on Arthur's Peak, Lory State Park, Ft. Collins.
At Wednesday's banquet, I'm afraid to say that Princeton once again dominated the dance floor as we all rocked out to the amazing Denver funk/rock/jam band Kinetix (great choice, Mike). The Stanford showing was disappointing in part because of the early departure of some of our most enthusiastic dancers. Don't get cocky though, Princeton. We'll be gunning for you next year.
The entirety of Tuesday morning's session was given over to communicating science. Dan Salkeld warmed up the crowd with some hilarious examples of the reporting frenzy that ensued following the publication of our paper on plague dynamics in prairie dog towns or, more recently, Hillary Young's work showing that excluding large ruminants increases rodent density in Kenya. Wow. Dan also used my Stanford colleagueRebecca Bird's work as an example of how an unexpected story can engage readers and listeners. My collaborator Tony Goldberg gave a talk that was also not lacking in ridiculous headlines thanks to his "viral" nose-tickwork. David Quammen, author of outstanding popular science books such as The Song of the Dodo and Spillover (which Bill Durham and I use for our class on environmental change and emerging infectious disease), gave a terrific presentation in which he consolidated a lot of nice, practical advice on the craft of writing engaging work into 18 points, amply illustrated by anecdotes of characters from our field. Sonia Altizer from the University of Georgia introduced the crowd to the opportunities (and pitfalls) of citizen science and suggested that it might just be possible to engage the public in disease ecology data collection. Some examples she identified included the granddaddy of citizen-science in the US run by the Laboratory of Ornithology at Cornell, the ZomBee Watch at SFSU, and her own Project MonarchHealth. If I had to summarize this session in one pithy phrase, I think it would have to be "Yay, ecologists!"
Quammen took to Twitter to call us out for being behind the curve with respect to social media.
@lfoquet I'll ask. But I think #EEID2014 is a quietish conference, not high on podcasts or social media. Tell me, others, if I'm wrong.
While there were, in fact, a few of us tweeting the occasional tidbit from the conference, I think his general point is valid. This stuff is intrinsically interesting and we can do a much better job communicating to broad publics.
Some talks that really caught my attention.
Ary Hoffmann gave a great talk about the complexities of using bacteria of the genus Wolbachia to control the Aedes mosquitoes that transmit dengue in Australia (and elsewhere). Wolbachia infects mosquitoes and can have a variety of effects on their biology. The reason artificial infection of mosquitoes wit this bacterium seems so promising as a means of biological control is that the offspring of crosses between infected and uninfected mosquitoes are not viable. This is obviously a very substantial fitness cost to the mosquitoes and this creates serious challenges for getting the infected mosquitoes to persist and take over local populations. Hoffmann presented a cool result about the invasibility of infected mosquitoes wherein in the early phases of introduction there is an unstable point in the mosquito dynamics. At this point, if the infected mosquitoes are above a threshold, they will successfully invade, otherwise, they will die out because of the inherent fitness costs of the Wolbachia infection. One policy challenge that arises is that to get a local population of mosquitoes above the invasibility threshold, researchers and vector-control specialists have to sometimes introduce a lot of mosquitoes. This means that the number of mosquitoes locally can increase substantially and, as you can imagine, this isn't always popular with communities.
Fellow Anthropologist Aaron Blackwell from UCSB gave a fantastic talk on our "old friends", the helminths (cue the freaky electron micrograph of a helminth's mouth!). Aaron participates in the Tsimane Health and Life History Project which was started by colleagues Mike Gurven (also at UCSB) and Hilly Kaplan (New Mexico). Using sophisticated multi-state Markov hazard models (go Anthropology!), Aaron showed that co-infection with helminths and Giardia is less frequent than expected among this population that experiences ubiquitous exposure to both pathogens and that, in fact, infection with the one appears to be protective against infection with the other. One of the most provocative results he presented showed that helminth infection actually lowered systolic blood pressure in men by an amount equivalent to the increase that comes from aging ten years. Chronic helminthic infection may be a reason why Tsimane men's systolic blood pressure does not rise precipitously with age as it does in the US. This result, which may provide fresh insights into the mechanisms of hypertension, a major source of morbidity in the US, struck me as particularly poignant given the demeaning comments made about NSF funding for work among the Tsimane from none other than Lamar Smith (R–TX), the chair of the House Committee on Science, Space, and Technology.
Anna Savage, a post-doc with the National Zoo in Washington DC, gave an awesome talk on the comparative immunogenetics of of frogs with respect to infection with the devastating fungal infection, chytridiomycosis. Chytridiomycosis has been identified as a major cause of amphibian extinction worldwide and Anna showed surprising heterogeneity in immune response across frog species. This is a subject with which I have only passing familiarity, but her talk demonstrated an amazing sophistication in integrating different levels of biological organization and making sense of a dauntingly complex problem. I would wager that Dr. Savage is one to keep an eye on.
The organizers tried a scheduling format that was a bit different from last year, wherein each session started with two half-hour talks generally given by somewhat more senior people. The second half of each session was then given over to brief ten-minute talks, typically delivered by more junior people. This format is nicely in keeping with the great EEID tradition of promoting the research of junior scientists. A few short talks that I found especially interesting included one by Sarah Hamer, from Texas A&M, on Chagas disease in the United States. She presented sobering data from national blood-bank surveillance showing a surprising number of Chagas-infected samples coming from donors with no history of travel to Latin America. When pushed by a questioner, she suggested that she would consider Chagas to be endemic in the US, at least in dogs and possibly even in people. Carrie Cizauskas, formerly of Wayne Getz's shop at Berkeley and now with Andy Dobson and Andrea Graham at Princeton, give a nice talk on the role of both stress and sex hormones in mediating macroparasite infection in wild ungulates in Etosha National Park, Namibia. Romain Garnier from Princeton described a very nifty image-processing approach to scanning large volumes of histological slides for indications of infection.
I perhaps didn't see as many posters as I should have. The problem with the poster sessions is that one keeps running into various people one wants to talk to. I did manage to check out the poster of my former freshman advisee and current Princeton EEB student Cara Brook. She's got an awesome PhD project studying the multi-host ecology of infectious disease in Malagasy fruit bats.
I'm looking forward to next year's meeting at the University of Georgia already. I'm also looking forward to resuscitating the pedagogical workshop that used to be a signature feature of this EEID meeting. More on that later...
In the spirit of my professed guilty amusement about the names of agricultural diseases, I just chuckled a bit at a promedmail update of what sounded like a biblical plague that had to be sent directly to the Apocrypha: Crayfish plague in Israel. Watch out, Pharaoh...
The mosquito, Aedes aegypti, which is the vector for a number of world scourges (e.g., dengue, yellow fever), has been found in San Mateo County (just across San Francisquito Creek from Stanford) for the first time since 1979. That makes three counties in California where the mosquito has been found. While not a panic-inducing development, it would be most excellent if the good people of San Mateo and Santa Clara counties would make sure their yards are free of mosquito breeding habitat!
I am recently back from the 2013 Ecology and Evolution of Infections Disease Conference at Penn State University. This was quite possibly the best meeting I have ever attended, not even for the science (which was nonetheless impeccable), but for the culture. I place the blame for this awesome culture firmly on the shoulders of the leaders of this field and, in particular, the primary motivating force behind the recent emergence of this field, Penn State's Peter Hudson. Since I had attended the other EEID conference at UGA earlier this Spring (another great conference), I had no intention on attending the Penn State conference this year. Then, one day in late March, Nita Bharti asked me if I was going and mentioned, "You know it's Pete's 60th birthday, right?" Well that sealed it; I really had no choice. I simply had to go if for no other reason than to pay my due respect to this man I admire so greatly. Pete has the most relentless optimism about the future of science and a willingness to make things happen that I have ever encountered and, in this way, has provided me one of my primary role models as a university professor and mentor. He has played a role in developing so many of the brilliant people who make this field so exciting, it's amazing (just a sample that comes immediately to mind: Ottar Bjornstad, Matt Ferrari, Nita Bharti, Marcel Salathé, Isabella Cattadori, Jamie Lloyd-Smith, Shweta Bansal, Jess Metcalf...). Of course, even as I write this, I realize the joint influence of another major player in the field, Bryan Grenfell, formerly of Penn State but now at Princeton, becomes obvious. A great scientist in his own right, Pete is the master facilitator, providing the support (and institutional interference!) that allows young scholars to thrive. He is a talent-spotter extraordinaire.
The talks that made up the bulk of the scientific program were, for the most part, excellent. The average age of the speakers was about 30, maybe just a bit higher. When one attends an academic conference, one typically expects that the major addresses to the collected masses will be by geezers, er, senior scholars in the field. There was a clear play at inversion of the standard model here though. Speakers were clearly chosen because of their trajectories, not their past achievements. That's pretty great. When I went up for tenure at Stanford, I was told that Stanford does not really care about what you have done; it cares about what you will do. Of course, the best information that the university has about your future work is the work you have already done. This conference embodied this spirit by placing the future (and, in many cases, current) leaders of the field in the key speaking roles while some of the biggest names in ecology, population biology, and epidemiology sat happily in the audience (e.g., joining Hudson and Grenfell were Andy Dobson, Andrew Read, Mick Crawley, Charles Godfray, Mike Boots, Mercedes Pascual, Les Real, Matt Thomas, ...)
The tone set by these great mentors carries through to the whole culture of the conference, where senior people attended the poster sessions, sat with students at lunches and dinners, and schmoozed at the plentiful open-bar mixers. For example, on the first full day of the conference, there was an afternoon poster session that started at 4:30 (we had been in back-to-back sessions since 8:30). This session was preceded by an hour-long poster-teaser session in which grad students and post-docs got up and presented 60-second (and, as Andrew Read noted, not one nanosecond more) teasers of their posters. Bear in mind, this session was entirely comprised of students and post-docs. It was striking that essentially every seat in the house was occupied and all the major players were present. The teasers were great – many were very funny, including a haiku apparently written by a triatomine bug and translated to us by Princeton EEB student Jennifer Peterson.
After the teasers, the conference went en masse to the fancy new Millenium Science Complex (it turns out that Pete Hudson has physical capital projects in addition to human capital ones!). There, participants milled about the 150 posters. After spending quite a bit of time doing this – and dutifully getting pictures of all my lab with their posters – I thought to check the time and realized it was nearly 6:30. The poster session had been going for two hours and nearly everyone was still there, including all the luminaries. It helped that there was free beer. I tweeted my amazement at this realization:
That is, in fact, Princeton's Bryan Grenfell moving fast in the middle of the picture, apparently making a bee-line for Michigan's Aaron King. Andrew Read is in the far background, talking to a poster-presenter (he has that posture).
Scientific highlights for me included Caroline Buckee's talk about measuring mobility in the context of malaria transmission in Kenya and Derek Cummings's talk on the Fluscape Project to measure spatial heterogeneity in influenza transmission in China. I am a long-time fan of this project and it's nice to see the great work that has come out of it. These talks were right in my wheelhouse of interest, but there were plenty other cool ones including Britt Koskella's talk on the dynamics of bacteria and phage on tree leaves.
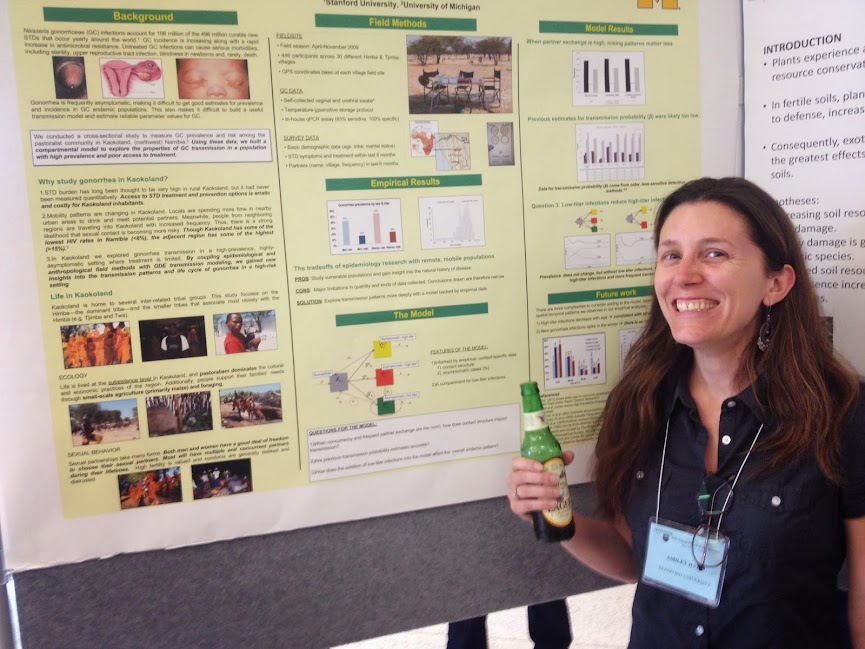
Stanford was exceedingly well represented at this conference. My lab had no fewer than five posters. Ashley Hazel presented on her work with Carl Simon on modeling gonorrhea transmission dynamics in Kaokoland, Namibia. Whitney Bagge presented her work on remote-sensing of rodent-borne disease in Kenya. Alejandro Feged presented work on the transmission dynamics of malaria in the Colombian Amazon among the indigenous Nukak people. Laura Bloomfield presented her remote sensing and spatial analysis work from our project on the spillover of primate retroviruses in Western Uganda. I closed things out with a minimalist poster on simple graphical models for multiple attractors in vector-borne disease dynamics in multi-host ecologies. In addition to my lab group, Giulio De Leo (with whom I have been running a weekly disease ecology workshop at Woods since winter quarter) was there, helping to bridge all sorts of structural holes in our collective collaboration graphs.
The other thing that comes out of these meetings, especially more intimate ones like EEID, is some actual work on collaborative projects. I managed to find some time to sit down and discuss plans with collaborators as well as do some shameless recruitment for my planned re-submission of the Stanford Biodemography Workshops. I'm really excited about some of these collaborations, including one that brings together my two major areas of interest: biodemography and life history theory and infectious disease ecology.
Oh, and I'm convinced that there must be an interpretive dance component to the Ph.D. exam in the Grenfell lab. This is certainly the most parsimonious explanation for much of what I saw Wednesday night.
notes on human ecology, population, and infectious disease

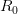
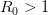
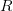
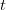
![log[Y(t)]/t](http://monkeysuncle.stanford.edu/wp-content/plugins/latex/cache/tex_380b1cea9aadb2220e7ff6b252ec1556.gif)
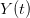
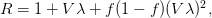
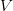
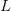
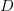
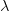
![\log[Y(t)]/t](http://monkeysuncle.stanford.edu/wp-content/plugins/latex/cache/tex_60ed99c4631e6be2f173613ae73e67f9.gif)
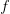
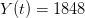


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}