Biological and Human Dimensions of Primate Retroviral Transmission
One of the great enduring mysteries in disease ecology is the timing of the AIDS pandemic. AIDS emerged as a clinical entity in the late 1970s, but HIV-1, the retrovirus that causes pandemic AIDS, entered the human population from wild primates many decades earlier, probably near the turn of the 20th century. Where was HIV during this long interval? We propose a novel ecological model for the delayed emergence of AIDS. Conceptually, in a metapopulation consisting of multiple, loosely interconnected sub-populations, a pathogen could persist at low levels indefinitely through a dynamic balance between localized transmission, localized extinction, and long-distance migration between sub-populations. This situation might accurately describe a network of villages in which population sizes are small and rates of migration are low, as would have been the case in Sub-Saharan Africa over a century ago.
We will test our model in a highly relevant non-human primate system. In 2009, we documented three simian retroviruses co-circulating in a metapopulation of wild red colobus monkeys (Procolobus rufomitratus) in Kibale National Park, Uganda, where we have conducted research for over two decades. We will collect detailed data on social interactions, demography, health, and infection from animals in a core social group.
We will also study a series of 20 red colobus sub-populations, each inhabiting a separate, isolated forest fragment. We will determine the historical connectivity of these sub-populations using a time series of remotely sensed images of forest cover going back to 1955, as well as using population genetic analyses of hypervariable nuclear DNA markers. We will assess the infection status of each animal over time and use viral molecular data to reconstruct transmission pathways.
Our transmission models will define the necessary conditions for a retrovirus to persist, but they will not be sufficient to explain why a retrovirus might emerge. This is because human social factors ultimately create the conditions that allow zoonotic diseases to be transmitted from animal reservoirs and to spread. We will therefore conduct an integrated analysis of the root eco-social drivers of human-primate contact and zoonotic transmission in this system. We will study social networks to understand how social resources structure key activities relevant to human-primate contact and zoonotic transmission risk, and we will explore knowledge, beliefs, and perceptions of human-primate contact and disease transmission for a broad sample of the population. We will reconcile perceived risk with actual risk through a linked human health survey and diagnostic testing for zoonotic primate retroviruses.
The ultimate product of our research will a data-driven set of transmission models to explain the long-term persistence of retroviruses within a metapopulation of hosts, as well as a linked analysis of how human social factors contribute to zoonotic infection risk in a relevant Sub-Saharan African population. Our study will elucidate not only the origins of HIV/AIDS, but also how early-stage zoonoses in general progress from "smoldering" subclinical infections to full-fledged pandemics.
I am thrilled to report that our latest EID project proposal, Biological and Human Dimensions of Primate Retroviral Transmission, has now been funded (by NIAID nonetheless!). I will briefly describe the project here and then shamelessly tack on the full text of our advertisement for a post-doc to work as the project manager with Tony Goldberg, PI for this grant, in the College of Veterinary Medicine, University of Wisconsin, Madison. This project will complement the ongoing work of the Kibale EcoHealth Project. The research team includes: Tony, Colin Chapman (McGill), Bill Switzer (CDC), Nelson Ting (Iowa), Mhairi Gibson (Bristol), Simon Frost (Cambridge), Jennifer Mason (Manchester), and me. This is a pretty great line-up of interdisciplinary scholars and I am honored to be included in the list.
Biological and Human Dimensions of Primate Retroviral Transmission
One of the great enduring mysteries in disease ecology is the timing of the AIDS pandemic. AIDS emerged as a clinical entity in the late 1970s, but HIV-1, the retrovirus that causes pandemic AIDS, entered the human population from wild primates many decades earlier, probably near the turn of the 20th century. Where was HIV during this long interval? We propose a novel ecological model for the delayed emergence of AIDS. Conceptually, in a metapopulation consisting of multiple, loosely interconnected sub-populations, a pathogen could persist at low levels indefinitely through a dynamic balance between localized transmission, localized extinction, and long-distance migration between sub-populations. This situation might accurately describe a network of villages in which population sizes are small and rates of migration are low, as would have been the case in Sub-Saharan Africa over a century ago.
We will test our model in a highly relevant non-human primate system. In 2009, we documented three simian retroviruses co-circulating in a metapopulation of wild red colobus monkeys (Procolobus rufomitratus) in Kibale National Park, Uganda, where we have conducted research for over two decades. We will collect detailed data on social interactions, demography, health, and infection from animals in a core social group.
We will also study a series of 20 red colobus sub-populations, each inhabiting a separate, isolated forest fragment. We will determine the historical connectivity of these sub-populations using a time series of remotely sensed images of forest cover going back to 1955, as well as using population genetic analyses of hypervariable nuclear DNA markers. We will assess the infection status of each animal over time and use viral molecular data to reconstruct transmission pathways.
Our transmission models will define the necessary conditions for a retrovirus to persist, but they will not be sufficient to explain why a retrovirus might emerge. This is because human social factors ultimately create the conditions that allow zoonotic diseases to be transmitted from animal reservoirs and to spread. We will therefore conduct an integrated analysis of the root eco-social drivers of human-primate contact and zoonotic transmission in this system. We will study social networks to understand how social resources structure key activities relevant to human-primate contact and zoonotic transmission risk, and we will explore knowledge, beliefs, and perceptions of human-primate contact and disease transmission for a broad sample of the population. We will reconcile perceived risk with actual risk through a linked human health survey and diagnostic testing for zoonotic primate retroviruses.
The ultimate product of our research will a data-driven set of transmission models to explain the long-term persistence of retroviruses within a metapopulation of hosts, as well as a linked analysis of how human social factors contribute to zoonotic infection risk in a relevant Sub-Saharan African population. Our study will elucidate not only the origins of HIV/AIDS, but also how early-stage zoonoses in general progress from "smoldering" subclinical infections to full-fledged pandemics.
Post Doctoral Opportunity
The Goldberg Lab at the University of Wisconsin-Madison invites applications for a post-doctoral researcher to study human social drivers of zoonotic disease in Sub-Saharan Africa. The post-doc will be an integral member of a new, international, NIH-funded project focused on the biological and human dimensions of primate infectious disease transmission in Uganda, including social drivers of human-primate contact and zoonotic transmission. This is a unique opportunity for a post-doctoral scholar with training in the social sciences to study human-wildlife conflict/contact and health and disease in a highly relevant ecological setting. The following criteria apply.
- Candidates must have completed or be near to completing a PhD in the social sciences, in a discipline such as anthropology, geography, sociology, behavioral epidemiology, or a relevant discipline within the public health fields.
- Candidates must have a demonstrated interest in health and infectious disease.
- Candidates must have prior field experience in Sub-Saharan Africa.
- Candidates must be willing to relocate to Madison, Wisconsin for three years.
- Candidates must be willing to spend substantial time abroad, including in Sub-Saharan Africa and at partner institutions in the United Kingdom.
- Candidates must have experience with collection and analysis of both quantitative and qualitative data. Familiarity with methods such as social network analysis, GIS, participatory methods, and survey design would be advantageous.
The successful candidate will help lead a dynamic international team of students and other post-docs in a multi-institutional, multidisciplinary project. Duties involve a flexible combination of fieldwork, analyses, and project coordination, in addition to helping to mentor students from North America, Europe, and Africa. The successful applicant will be expected to explore new research directions of her/his choosing, assisted by a strong team of collaborators.
University of Wisconsin-Madison is a top-notch institution for research and training in the social and health sciences. Madison, WI, is a vibrant city with outstanding culture and exceptional opportunities for outdoor recreation.
Applicants should send a current CV, a statement of research interests and qualifications (be sure to address the six criteria above), and a list of three people (names, addresses, e-mails) who can serve as references.
Materials and inquiries should be sent to Dr. Tony L. Goldberg (tgoldberg@vetmed.wisc.edu). Application materials must be received by September 12, 2011 for full consideration; the position is available starting immediately and requires a three-year commitment.
![log[Y(t)]/t](http://monkeysuncle.stanford.edu/wp-content/plugins/latex/cache/tex_380b1cea9aadb2220e7ff6b252ec1556.gif)
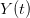
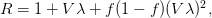
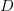
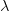
![\log[Y(t)]/t](http://monkeysuncle.stanford.edu/wp-content/plugins/latex/cache/tex_60ed99c4631e6be2f173613ae73e67f9.gif)
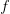


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}