It's been a long time since I've written in monkey's uncle. Life has gotten pretty busy and my seeming inability to write brief entries has led me to neglect the blog this year. However, I am freshly back from the Ecology and Evolution of Infectious Disease Conference in Fort Collins, Colorado and feel compelled to give my annual run-down. The conference was hosted by friend and colleague Mike Antolin, Sue Vandewoude, and my erstwhile post-doc, now CSU researcher, Dan Salkeld. Nice job, folks, on a very successful conference.
EEID is pretty much the best meeting. As I noted in last year's post, I love its future-orientation. EEID is a meeting that foregrounds the work of junior scientists and there was, as ever, a tremendous array of human capital on display at this meeting. This drives home to me the importance of investment in professional training and research programs that specifically develop human capital. This community exists in large measure because of the innovative program jointly offered by NSF and NIH. Thanks as ever to the vision and hard work of Josh Rosenthal, Sam Scheiner, and all the funders (e.g., support from The Gates Foundation can be found all around this conference) for this area. It's always great to catch up with smart, fun friends. Plenty of time was spent talking science and drinking craft beer (what a beer town Ft. Collins is!) with the likes of Peter Hudson, Jessica Metcalf, Ottar Bjornstad, Aaron King, Mike Antolin, Tony Goldberg, Issa Cattadori, Maciej Boni, Marm Kilpatrick and, of course, Dan Salkeld. It was nice to meet and chat, if only briefly, with my sometime remote collaborator Paul Sharp, who gave what I understand to be an extremely stimulating keynote on the complicated and surprising evolution of malaria (alas, I missed it as I was delayed getting to Ft. Collins). I also spent some quality time learning about acquired immunity in dogs with Colin Parrish. This may come in handy for some ideas that Jess Metcalf and I have been playing around with.
There is a great tradition of the EEID hike and closing banquet/dance. Ft. Collins provided a beautiful and challenging hike out in Lory State Park. The view from the top of Arthur's Peak was pretty amazing.

At Wednesday's banquet, I'm afraid to say that Princeton once again dominated the dance floor as we all rocked out to the amazing Denver funk/rock/jam band Kinetix (great choice, Mike). The Stanford showing was disappointing in part because of the early departure of some of our most enthusiastic dancers. Don't get cocky though, Princeton. We'll be gunning for you next year.
The entirety of Tuesday morning's session was given over to communicating science. Dan Salkeld warmed up the crowd with some hilarious examples of the reporting frenzy that ensued following the publication of our paper on plague dynamics in prairie dog towns or, more recently, Hillary Young's work showing that excluding large ruminants increases rodent density in Kenya. Wow. Dan also used my Stanford colleague Rebecca Bird's work as an example of how an unexpected story can engage readers and listeners. My collaborator Tony Goldberg gave a talk that was also not lacking in ridiculous headlines thanks to his "viral" nose-tick work. David Quammen, author of outstanding popular science books such as The Song of the Dodo and Spillover (which Bill Durham and I use for our class on environmental change and emerging infectious disease), gave a terrific presentation in which he consolidated a lot of nice, practical advice on the craft of writing engaging work into 18 points, amply illustrated by anecdotes of characters from our field. Sonia Altizer from the University of Georgia introduced the crowd to the opportunities (and pitfalls) of citizen science and suggested that it might just be possible to engage the public in disease ecology data collection. Some examples she identified included the granddaddy of citizen-science in the US run by the Laboratory of Ornithology at Cornell, the ZomBee Watch at SFSU, and her own Project MonarchHealth. If I had to summarize this session in one pithy phrase, I think it would have to be "Yay, ecologists!"
Quammen took to Twitter to call us out for being behind the curve with respect to social media.
@lfoquet I'll ask. But I think #EEID2014 is a quietish conference, not high on podcasts or social media. Tell me, others, if I'm wrong.
— David Quammen (@DavidQuammen) June 2, 2014
While there were, in fact, a few of us tweeting the occasional tidbit from the conference, I think his general point is valid. This stuff is intrinsically interesting and we can do a much better job communicating to broad publics.
Some talks that really caught my attention.
Ary Hoffmann gave a great talk about the complexities of using bacteria of the genus Wolbachia to control the Aedes mosquitoes that transmit dengue in Australia (and elsewhere). Wolbachia infects mosquitoes and can have a variety of effects on their biology. The reason artificial infection of mosquitoes wit this bacterium seems so promising as a means of biological control is that the offspring of crosses between infected and uninfected mosquitoes are not viable. This is obviously a very substantial fitness cost to the mosquitoes and this creates serious challenges for getting the infected mosquitoes to persist and take over local populations. Hoffmann presented a cool result about the invasibility of infected mosquitoes wherein in the early phases of introduction there is an unstable point in the mosquito dynamics. At this point, if the infected mosquitoes are above a threshold, they will successfully invade, otherwise, they will die out because of the inherent fitness costs of the Wolbachia infection. One policy challenge that arises is that to get a local population of mosquitoes above the invasibility threshold, researchers and vector-control specialists have to sometimes introduce a lot of mosquitoes. This means that the number of mosquitoes locally can increase substantially and, as you can imagine, this isn't always popular with communities.
Fellow Anthropologist Aaron Blackwell from UCSB gave a fantastic talk on our "old friends", the helminths (cue the freaky electron micrograph of a helminth's mouth!). Aaron participates in the Tsimane Health and Life History Project which was started by colleagues Mike Gurven (also at UCSB) and Hilly Kaplan (New Mexico). Using sophisticated multi-state Markov hazard models (go Anthropology!), Aaron showed that co-infection with helminths and Giardia is less frequent than expected among this population that experiences ubiquitous exposure to both pathogens and that, in fact, infection with the one appears to be protective against infection with the other. One of the most provocative results he presented showed that helminth infection actually lowered systolic blood pressure in men by an amount equivalent to the increase that comes from aging ten years. Chronic helminthic infection may be a reason why Tsimane men's systolic blood pressure does not rise precipitously with age as it does in the US. This result, which may provide fresh insights into the mechanisms of hypertension, a major source of morbidity in the US, struck me as particularly poignant given the demeaning comments made about NSF funding for work among the Tsimane from none other than Lamar Smith (R–TX), the chair of the House Committee on Science, Space, and Technology.
Anna Savage, a post-doc with the National Zoo in Washington DC, gave an awesome talk on the comparative immunogenetics of of frogs with respect to infection with the devastating fungal infection, chytridiomycosis. Chytridiomycosis has been identified as a major cause of amphibian extinction worldwide and Anna showed surprising heterogeneity in immune response across frog species. This is a subject with which I have only passing familiarity, but her talk demonstrated an amazing sophistication in integrating different levels of biological organization and making sense of a dauntingly complex problem. I would wager that Dr. Savage is one to keep an eye on.
The organizers tried a scheduling format that was a bit different from last year, wherein each session started with two half-hour talks generally given by somewhat more senior people. The second half of each session was then given over to brief ten-minute talks, typically delivered by more junior people. This format is nicely in keeping with the great EEID tradition of promoting the research of junior scientists. A few short talks that I found especially interesting included one by Sarah Hamer, from Texas A&M, on Chagas disease in the United States. She presented sobering data from national blood-bank surveillance showing a surprising number of Chagas-infected samples coming from donors with no history of travel to Latin America. When pushed by a questioner, she suggested that she would consider Chagas to be endemic in the US, at least in dogs and possibly even in people. Carrie Cizauskas, formerly of Wayne Getz's shop at Berkeley and now with Andy Dobson and Andrea Graham at Princeton, give a nice talk on the role of both stress and sex hormones in mediating macroparasite infection in wild ungulates in Etosha National Park, Namibia. Romain Garnier from Princeton described a very nifty image-processing approach to scanning large volumes of histological slides for indications of infection.
I perhaps didn't see as many posters as I should have. The problem with the poster sessions is that one keeps running into various people one wants to talk to. I did manage to check out the poster of my former freshman advisee and current Princeton EEB student Cara Brook. She's got an awesome PhD project studying the multi-host ecology of infectious disease in Malagasy fruit bats.
I'm looking forward to next year's meeting at the University of Georgia already. I'm also looking forward to resuscitating the pedagogical workshop that used to be a signature feature of this EEID meeting. More on that later...



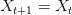

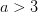




{kind=link}
{kind=link}