The logistic equation is a model of population growth where the size of the population exerts negative feedback on its growth rate. As population size increases, the rate of increase declines, leading eventually to an equilibrium population size known as the carrying capacity. The time course of this model is the familiar S-shaped growth that is generally associated with resource limitation. This model has only two parameters:  is the intrinsic growth rate and
is the intrinsic growth rate and 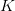 is the carrying capacity. The rate of increase in the population declines as a linear function of population size. In symbols:
is the carrying capacity. The rate of increase in the population declines as a linear function of population size. In symbols:

When the population size is very small (i.e., when 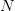 is close to zero), the term in the parentheses is approximately one and population growth is approximately exponential. When population size is close to the carrying capacity (i.e.,
is close to zero), the term in the parentheses is approximately one and population growth is approximately exponential. When population size is close to the carrying capacity (i.e., 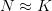 ), the term in parentheses approaches zero, and population growth ceases. It is straightforward to integrate this equation by partial fractions and show that resulting solution is indeed an S-shaped, or sigmoid, curve.
), the term in parentheses approaches zero, and population growth ceases. It is straightforward to integrate this equation by partial fractions and show that resulting solution is indeed an S-shaped, or sigmoid, curve.
Raymond Pearl was a luminary in human biology. A professor at Johns Hopkins University, a founder of the Society for Human Biology and the International Union for the Scientific Study of Population (IUSSP), Pearl also re-discovered the logistic growth model (which was originally developed by the great Belgian mathematician Pierre François Verhulst). In the logistic model, Pearl believed he had found a universal law of biological growth at its various levels of organization. In his book, The Biology of Population Growth, Pearl wrote:
... human populations grow according to the same law as do the experimental populations of lower organisms, and in turn as do individual plants and animals in body size. This is demonstrated in two ways: first by showing as was done in my former book "Studies in Human Biology," that in a great variety of countries all of the recorded census history which exists is accurately described by the same general mathematical equation as that which describes the growth of experimental populations; second, by bringing forward in the present book the case of a human population-the indigenous native population of Algeria-which has in the 75 years of its recorded census history practically completed a single cycle of growth along the logistic curve.
In addition to Algeria, Pearl fit the logistic model to the population of the United States from 1790-1930. The fit he produced was uncanny and he confidently predicted that the US population would level out at 198 million, since this was the best-fit value of in his analysis. I have plotted the US population size (from the decennial census) as black points below, with Pearl's fitted curve in grey. We can see that the curve fits incredibly well for the period 1790-1930 (the span to which he fit the data), but the difference between prediction and empirical reality becomes increasingly large after 1950 (yep, that would be thanks to the Baby Boom).

Why does the logistic model fail so spectacularly in this case (and many others)?
The logistic model is phenomenological, rather than mechanistic. A phenomenological model is a mathematical convenience that we use to describe some empirical observations, but has no foundations in mechanisms or first principles. Such models can be useful when theory is lacking to explain some phenomenon or when the mathematics that would be required to model the mechanisms is too complicated. You can make a prediction from a phenomenological model, but I wouldn't bet the farm on that prediction. In the absence of an actual understanding of the mechanisms producing the population change, the predictions can go horribly wrong, as we see in the case of Raymond Pearl's fit.
Specifically, the logistic model fails to consider mechanisms of population regulation. When density increases, what is affected? Birth rates? Death rates? The parameter in the logistic model is simply the difference in the gross birth and death rates when there are no conspecifics present. In general, when the birth rate exceeds the death rate, a population increases. The linear decrease in with increasing population size presumably can come about by either the birth rate decreasing or the death rate increasing. The logistic model is indifferent to the specific cause of slowing. It just stops increasing when 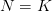 . Is it possible that, in real populations, increasing the death rate and decreasing the birth rate might have qualitatively different effects on population growth? We'll see.
. Is it possible that, in real populations, increasing the death rate and decreasing the birth rate might have qualitatively different effects on population growth? We'll see.
This probably goes without saying, but there is no capacity for the positive feedbacks with population size. In her classic work, The Conditions of Agricultural Growth, Danish economist Esther Boserup noted that population growth often stimulates innovation. Population pressure might cause an agricultural group that has run out of land to intensify cultivation by improving the land or multi-cropping, thereby facilitating even greater population growth. Various authors, including Ken Wachter and Ron Lee (both at Berkeley) and Jim Wood at Penn State have noted that real populations probably incorporate both Malthusian (i.e., conditions leading to increased mortality, decreased fertility, and general misery with increased population size) and Boserupian phases in their dynamics. Wood coined the term "MaB Ratchet" (MaB = Malthus and Boserup) which describes the following dynamic: Malthusian pressure incites Boserupian innovation, relaxing negative feedback and allowing further population growth. While a population is undergoing a Boserupian expansion, quality of life improves. Alas, given enough time, the population will always return to "the same level of marginal immiseration." (Wood 1998: 114). Such complex regimes of positive and negative population feedback are not a possibility .
One final problem with the logistic model is that there is no structure -- all individuals are identical in terms of their effect on and contribution to population growth. Human vital rates vary predictably – and substantially – by age, sex, geographic region, urban vs. rural residence, etc. And then there's the issue of unequal resource distribution. All individuals in a population are hardly equal in their consumption (or production) and so we should hardly expect each to exert an identical force on population growth.
So are there better alternative models for human population growth that incorporate the sensible idea that as populations push the limits of their resource base, growth should slow down and eventually cease? There is now. My Stanford colleague and collaborator in various endeavors, Shripad Tuljapurkar, has a series of papers in which he and his students develop mechanistic population models for agricultural populations that specifically link age-specific vital rates (i.e., survivorship, fertility), agricultural production and labor, and specific (age-specific) metabolic needs for individuals engaged in heavy physical labor. The models start with an optimal energy supply for survival and reproduction. As food gets more scarce, mortality increases and fertility decreases. The model has an equilibrium where birth and death rates balance. A key feature of the model is the idea of the food ratio, which is the number of calories available to consume in a given year relative to the number of calories needed to maximize survival and fertility. The food ratio tells us how hungry the population is. In the first of a series of three papers, Lee and Tuljapurkar (2008) develop this model and show how changes in mortality, fertility, and agricultural productivity actually all have distinct effects on the population growth rate, equilibrium, and how hungry people are at equilibrium. Analysis of their model yielded the following results:
- Increasing agricultural productivity or the amount of time spent working on agricultural production increases the food ratio, while keeping the population growth rate largely unchanged
- Increasing baseline survival increases the food ratio but decreases the population growth rate
- Decreasing fertility only decreases the growth rate – the food ratio remains unchanged
So, we see that it is possible that increasing the death rate and decreasing the birth rate might have qualitatively different effects on population growth. In fact, it seems quite likely, given Lee & Tulja's model.
We don't, as yet, have the kind of test that we gave Raymond Pearl's application of the logistic model to US population size. It would be very nice if we could use the Lee-Tulja model to make a prediction about the future dynamics of some population (and its distribution of hunger) and challenge this prediction with data not used for fitting the model in the first place. This said, I think that theoretical exercise alone is enough to demonstrate the importance of moving beyond phenomenological population models whenever possible. We are unlikely to make accurate predictions or understand the response of population to environmental and social changes in the absence of mechanistic models.
References
Lee, C. T., and S. Tuljapurkar. 2008. Population and prehistory I: Food-dependent population growth in constant environments. Theoretical Population Biology. 73:473–482.
Wood, J. W. 1998. A theory of preindustrial population dynamics: Demography, economy, and well-being in Malthusian systems. Current Anthropology. 39 (1):99-135.