I am currently teaching a class entitled "Demography and Life History Theory." For this class, we read the classic paper by Madhav Gadgil and Bill Bossert, "Life Historical Consequences of Natural Selection." In preparing for class, I re-read this paper for about the twelfth time. Something happened this time. It really dawned on me what a spectacularly important paper this is. Just about every important theme in life history theory is addressed in this paper and the analyses remain remarkably relevant.
One of the fundamental ideas this paper brought to life history theory is thinking about the convexity of the functions that describe both the fitness benefits and costs associated with the degree of reproductive effort. In particular, Gadgil & Bossert show that iteroparity (i.e., repeated breeding) can only evolve if the function relating benefit to effort is concave or the function relating cost to effort is convex. The figure shows a concave profit function and a linear cost. Clearly, the maximal value of the difference between the profit and cost happens at some intermediate level of effort 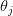 .
.

Maddeningly, Gadgil & Bossert invert the terms "convex" and "concave." I'm sure there is a good historical explanation for this, but contemporary usage indicates that a continuous, twice differentiable function 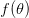 is convex if
is convex if  . That is a convex function shows increasing marginal returns to effort, whereas a concave function shows diminishing marginal returns to effort.
. That is a convex function shows increasing marginal returns to effort, whereas a concave function shows diminishing marginal returns to effort.
Their analysis focuses on a discrete-time form of Lotka's characterisitc equation:
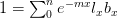
where 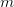 is the Maulthusian parameter (the intrinsic rate of increase,
is the Maulthusian parameter (the intrinsic rate of increase,  , in a density-independent population),
, in a density-independent population), 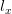 is the fraction surviving to age
is the fraction surviving to age  ,
, 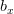 is the birth rate (in daughters) to females age .
is the birth rate (in daughters) to females age .
What makes this approach so interesting and important is that the vital rates are functions of reproductive effort 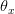 at each age. In addition to and being functions of effort, they are also functions of "satisfaction"
at each age. In addition to and being functions of effort, they are also functions of "satisfaction" 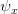 , a measure of environmental quality. Fertility at age increases with effort and satisfaction. Survivorship and growth decrease with effort and increase with satisfaction. Fertility also increases with body size.
, a measure of environmental quality. Fertility at age increases with effort and satisfaction. Survivorship and growth decrease with effort and increase with satisfaction. Fertility also increases with body size.
Gadgil and Bossert then maximize subject to the biological constraints on intrinsic mortality, growth rate, and initial size, and the environmental constraints of satisfaction and mortality due to predation. They used an automatic computer to numerically solve their optimization problem. They derive a number of quite general results for age-structured populations.
- Only when the profit function is concave or the cost function is convex can repeated breeding be optimal.
- Reproductive effort increases with age in repeated breeders. George Williams arrived at a similar conclusion in 1966, but failed to consider the possibility that the fitness profit could decline with age. Gadgil & Bossert arrive at this result with a much more general approach than that used by Williams.
- When mortality increases following some age
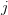 , reproductive effort increases for ages less than .
, reproductive effort increases for ages less than . - When reproductive potential increases slowly with size, reproductive effort will be lower at maturity, rise with age, and growth will continue beyond maturity.
- A uniform change in mortality -- affecting all ages equally -- will have no direct impact on reproductive effort.
- If the population is resource-limited, such a uniform change in mortality will increase satisfaction
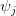 with its consequent effects, including a lowering of age at first breeding and increase in reproductive effort.
with its consequent effects, including a lowering of age at first breeding and increase in reproductive effort.
I would have to say that this is the desert island paper for life history theory. If you only ever read one paper (conditional on having the mathematics background to make it worthwhile), this is the one to read, even after 38 years.