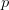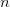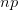Yesterday, I spent most of the day collecting content for my upcoming classes this spring and getting the course web sites together. For the first time in a while, I will (officially) be teaching two classes in one quarter (which effectively means teaching three or four when I add the other things like lab meetings in). The first is our graduate class on statistics in the anthropological sciences. I taught something like this back in the old department (i.e., Anthropological Sciences) but haven't taught it in years (though a Google search for "department of anthropological sciences stanford" turns up the syllabus for this class). It is technically a requirement for Ph.D. students in the Ecology and Environment focus within Anthropology, so it's about time. It will be fun to teach again and we're looking to use the class as a platform to develop resources for anthropologists doing statistical work (more later).
The other class that I will be teaching starting next week is Ecology, Evolution, and Human Health, a class I first taught last year. This class is meant to be an introduction to the Ecology and Environment undergraduate focus in Anthropology. I'm actually really looking forward to teaching it again. The course material forms the core of a book I am writing on human population biology and my attempts at improving the lectures has done wonders for my writing output of late. We'll see what happens when the quarter actually starts. Hopefully, between trips to Rwanda and Tanzania and moving into Arroyo House this summer, I will find time to finish it!
Back in December, when the is-anthropology-science kerfuffle was going strong, I wrote a blog post in which I suggested that if you want to feel good about the future of scientific anthropology (which, I admit, can sometimes be difficult, even for an obstinate optimist), all you need to do is look at the great work coming from the new generation of trans-disciplinary anthropologists (and other biosocial scientists). At the time, I put together a short list of people whose work I greatly admire. These included:
- Craig Hadley at Emory on food security and psychological well-being
- Amber Wutich at ASU on vulnerability, water security, and common-pool resources
- Lance Gravlee at UF on the embodiment of racial discrimination and its manifestations in health
- Brooke Scelza at UCLA on parental investment and childhood outcomes
- Dan Hrushka at ASU on how cultural beliefs, norms and values interact with economic constraints to produce health outcomes
- Crickette Sanz at Washington University on multi-ape ecology of the Goualougo Triangle, Republic of Congo
- Herman Pontzer at CUNY on measuring daily energy expenditures in hunter-gatherers
- Rebecca and Douglas Bird on subsistence and signaling among Martu foragers
In preparing for Anthro 31, I started to put together a list of links to people doing the kind of work we will discuss. In a pique of obsessiveness yesterday, I greatly expanded that list. It occurred to me that this list is somewhat orphaned in an obscure directory for a particular class I occasionally teach and that it would make sense to share it more generally. So, here we go, copied wholesale from my class links page (though that page still contains links to books, professional societies, and other resources for students interested in human ecology, demography, health, etc.):
There are a number of excellent practicing anthropologists who maintain science blogs. Among these are Kate Clancy's (UIUC) Context and Variation, Daniel Lende and Greg Downey's Neuroanthropology, Julienne Rutherford's AAPA BANDIT, and Patrick Clarkin's blog dedicated to biological anthropology, war and health, growth nutrition. Along with Rebecca Stumpf, Kate Clancy is also the director of the Laboratory for Evolutionary Endocrinology (which has its own blog) at the University of Illinois.
Upon further reflection, I think that the University of Illinois has to be a major contender for best place to study biological anthropology. Wow, they've got an amazing group of biological anthropologists there. Stanley Ambrose, Kate Clancy, Paul Garber, Lyle Konigsberg, Steve Leigh, Ripan Malhi, John Polk, Charles Roseman, Laura Shackelford, Rebecca Stumpf. Too many to link to directly. I don't know all of them, but the ones I know are outstanding. Yipes! I think they may be plotting to take over the field.
Back to the blog front, you can always count on gems of anthropological, evolutionary, and political wisdom from Greg Laden as well.
Susan C. Antón (NYU) and Josh Snodgrass (Oregon) organize the Bones and Behavior Working Group, the goal of which is to foster greater synthesis across the different sub-areas of biological anthropology. Of particular interest are their standardized protocols for anthropometry.
Mario Luis Small, at the University of Chicago, has done some really outstanding work measuring how social institutions affect social capital and the impact such differences in social capital actually have for people's well-being.
Richard Bribiescas is the author of Men: Evolutionary and Life History and is director of the Reproductive Ecology Laboratory at Yale. Yale is also now the home to Catherine Panter-Brick who also happens to be the senior editor for medical anthropology at Social Science and Medicine.
A number of excellent human biologists find their home in the Laboratory for Human Biology Research at Northwestern. This includes Bill Leonard, Thom McDade, and Chris Kuzawa. Rumor has it that alumna Elizabeth Sweet is moving back to Northwestern as well. She is doing truly innovative work integrating the rigorous analysis of biomarkers of health (and a bicultural perspective favored by the Northwestern group) and the political economy of economic and social disparities -- really getting at how inequality 'gets under the skin.' I really look forward to seeing what comes from her future research.
Karen Kramer, in the department formerly known as (Biological) Anthropology at Harvard, is a real leader in integrating evolutionary, demographic, and economic perspectives on human reproduction and the life histories.
Patrick Clarkin at UMass, Boston has a very interesting research program employing biocultural and evolutionary models to understand the effects of war on nutrition and growth among SE Asian diaspora. UMass, Boston is also home to Colleen Nyberg who does great work on acculturation and health, the psychobiology of stress and HPA function, and growth and development.
Julienne Rutherford at the University of Illinois, Chicago School of Dentistry works on the role of the intrauterine environment on health. Of particular interest for this class is her collaborative work on understanding the epigenetic regulation of placental systems of amino acid transport as part of the Cebu Longitudinal Study in the Philippines. UIC also has a number of excellent human biologists scattered about in anthropology, including Betsy Abrams and Crystal Patil, Epidemiology (Bob Bailey) and Community Health Sciences (Nadine Peacock).
Let's not forget our friends across The Pond. Durham may have lost Catherine Panter-Brick to Yale, but they got a number of new folks who, when combined with the veterans, make it a very appealing place to study ecological/evolutionary anthropology. Among the faculty there are my colleagues Gillian Bentley, Rebecca Sear, and Frank Marlowe, and numerous others. Rebecca does very sophisticated work in anthropological demography, while Frank is one of the leading ethnographers of contemporary hunter-gatherers (and my collaborator on our Hadza demography project).
Ruth Mace, in my opinion, does some of the best work in human behavioral ecology right now and she keeps churning out top students at UCL.
I'm looking forward to working with Mhairi Gibson at Bristol on our new project on the transmission dynamics of primate retroviruses and human-wildlife contact in Uganda. She has done excellent work on the behavioral ecology of reproduction and parental investment in Ethiopia.
I will also mention a number of excellent researchers who teach classes that are relevant to Ecology, Evolution, and Human Health:
Mark Moritz at Ohio State University has established a Hunter-Gatherer Wiki is conjunction with his course on Hunter-Gatherers. Mark came and gave a terrific talk on livestock exchanges among FulBe pastoralists at the MAPSS colloquium this year.
Mike Gurven at UCSB teaches a course on the behavioral ecology of hunter-gatherers. Mike does some of the most interesting biodemographic work out there these days.
Bruce Winterhalder at UC Davis, a founding father of human behavioral ecology, has a very interesting course on classics in cultural ecology.
Claudia Valeggia, at Penn, does great work among the Toba people of Argentina teaches a class on reproductive ecology.
Lots of good people. Lots of good work. Surely, there is reason for optimism...